Для корневых деревьев используем аналогичную концепцию. Два корневых дерева топологически эквивалентны, если одно можно преобразовать в другое, не перемещая корень. Можно изменить длину ребер, но не структуру ветвления.
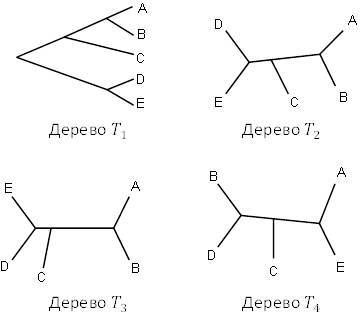
Рисунок 5.3. Четыре топологических дерева; как некорневые деревья, все, кроме правого нижнего, они идентичны.
Вопросы для самопроверки:
– Как на рисунке 5.3 расположить корень дерева
, чтобы полученное дерево не было топологически эквивалентным корневому дереву
? А чтобы получилось топологически то же самое, что и корневое дерево
?
Топологическое дерево, даже некорневое, довольно многое говорит об эволюционной истории таксонов, к которым оно относится. Например, все деревья на рисунке 5.2 показывают, что таксоны
и
связаны одним разделением линии, точно так же как
и
. Тем не менее, несколько раздвоений линии произошли между
и
, эволюционировавших от общего предка, поскольку в процессе возникли два других таксона.
Знание местоположения корня передает больше информации и может дать лучшее представление о порядке событий во времени. Например, изображенное справа на рисунке 5.2 дерево однозначно задаёт следующий порядок бифуркаций: общий предок дал начало двум таксонам, один из которых, возможно, эволюционировал дальше, чтобы стать
; другой впоследствии породил
и третий таксон; этот третий таксон затем породил
и
.
Дерево в центре рисунка 5.2 можно интерпретировать аналогичным образом. Общий предок дал начало двум таксонам, один из которых дал начало как
, так и
, в то время как другой дал начало
и
. Обратите внимание, однако, что только с топологическим деревом не можем сказать, какое из этих двух последних бифуркации произошло первым: существовал ли самый последний общий предок
и
более поздний, чем
и
? Нет возможности определить это по дереву.
Количество различных топологических деревьев, которые могут соотносить несколько терминальных таксонов, быстро растёт с увеличением числа таксонов. Например, существует только 1 некорневое топологическое дерево, относящееся к 3 таксонам, но есть 3 некорневых топологически различных дерева, относящиеся к 4 таксонам.
Вопросы для самопроверки:
– Нарисуйте одно некорневое топологическое дерево, которое может относиться к терминальным таксонам
,
и
. Нарисуйте три некорневых топологических дерева, которые могут относиться к терминальным таксонам
,
,
и
.
На 5 терминальных таксонов приходится 15 таких деревьев. Таким образом, если не принимать во внимание местонахождение корня, существует на 13 деревьев, которые могут связать 5 гоминоидов, а больше, чем было представлено во введении к главе. Для 6 терминальных таксонов насчитывается более 100 возможных некорневых деревьев. По мере увеличения числа таксонов количество деревьев быстро вырастает до астрономических размеров. В упражнениях найдете точные формулы, определяющие количество некорневых и корневых деревьев, относящихся к
таксонам. Также увидите, насколько велики эти числа, даже для относительно небольшого числа таксонов. Большое количество деревьев вызывает дискомфорт, потому что это означает, что некоторые подходы к поиску хорошего дерева для соотнесения таксонов будут медленными. Если метод находит «лучшее» дерево, рассматривая каждое возможное дерево по отдельности, то его использование будет чрезвычайно трудоемким, когда задействовано много таксонов.
На помощь в решении обозначенной проблемы поиска лучшей классификации приходят метрические деревья. В дополнение к топологической структуре дерево может иметь метрическую структуру; каждому ребру может быть присвоена определенная длина. Эта метрическая структура может быть задана путем записи чисел для обозначения длин рядом с ребрами (см. Рисунок 5.4 (слева)), или ребро может быть наглядно представлено путем рисования дерева с ребрами соответствующей длины, но без их явной нумерации. Таким образом, топологическое дерево и немаркированное метрическое дерево неотличимы друг от друга. Для ясности, будем маркировать ребра их длиной, когда нужно задать метрическое дерево.
Как правило, длины ребер в филогенетическом дереве, построенном из данных последовательности ДНК, каким-то образом представляют собой количество мутаций, которые произошли между расщеплениями линии. Чем длиннее ребро, тем больше последовательность ДНК мутировала в ходе эволюции, которую представляет это ребро.
Если, например, модель Джукса-Кантора замещения оснований адекватно описала эволюцию нескольких таксонов, то длина ребра в дереве, относящемся к ним, может быть расстоянием Джукса-Кантора между последовательностями на двух концах. Как видели в главе 4, это расстояние представляет собой среднее число замен оснований на сайт, произошедших при происхождении новой последовательности. Сюда включены мутации, скрытые другими мутациями, для оценки которых была разработана формула расстояния. Поскольку расстояние Джукса-Кантора является аддитивным и симметричным, общее расстояние между двумя таксонами вдоль дерева должно быть расстоянием Джукса-Кантора между ними.
Если предположение о молекулярных часах справедливо для эволюции связанных последовательностей, то расстояния в дереве имеют постоянное значение. Напомним, что молекулярные часы просто означают, что скорость мутаций постоянна для всех рассматриваемых линий. Если
обозначает скорость мутации, измеряемую, например, в количестве произошедших за год замен оснований на сайт, а
обозначает время в годах, то количество мутаций, которое произойдет в течение этого времени, составляет
базовых замен на сайт.