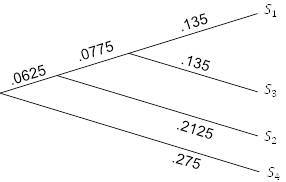
Снова объединив таксоны, формируем группу
и вычисляем расстояние от неё до
путем усреднения исходных расстояний от
до каждого из
,
и
. Это приводит к значению
. Обратите внимание, что это не то же самое, что усреднение расстояния от
до
и до
. Поскольку новая таблица расстояний будет иметь это значение в качестве единственной записи, нет необходимости приводить ее. Изобразим рисунок 5.8, считая, что расстояние от корня до
равно
. Конечное ребро имеет длину. 0625, таким образом, помещаем оставшийся таксон на расстоянии
от корня.
Рисунок 5.8. UPGMA; шаг 3.
Как и подозревали, дерево, которое построили для имеющихся данных, не совсем соответствует этим данным. Расстояние на дереве от
до
, например, равно
, хотя по исходным данным должно быть
. Тем не менее, расстояния между вершинами построенного дерева, по крайней мере, достаточно близки к расстояниям, указанным в исходных табличных данных.
Если бы было больше таксонов, то пришлось бы сделать больше шагов для завершения процесса UPGMA, но не было бы никаких принципиально новых действий. На каждом шаге объединяем два ближайших таксона или группы вместе, всегда размещая их на равных расстояниях от общего предка. Затем сворачиваем объединенные таксоны в группу, используя усреднение для вычисления расстояния от этой группы до таксонов и групп, которые еще предстоит объединить. Один момент, с которым следует быть особенно осторожным, заключается в том, что при вычислении расстояний между двумя группами нужно усреднить все расстояния от членов одной группы до членов другой – если одна группа имеет
членов, а другая имеет
членов, придется усреднить
расстояний. Каждый шаг алгоритма уменьшает размер таблицы расстояний на единицу, так что после достаточного количества шагов все таксоны объединяются в единое дерево.
Обратите внимание, что предположение о молекулярных часах неявно присутствовала в UPGMA. В примере, когда поместили
и
на концы ветвей одинаковой длины, предположили, что количество мутаций, которые каждый из них претерпел от своего общего предка, было одинаковым. Метод UPGMA всегда размещает все таксоны на одинаковом расстоянии от корня, так что количество мутаций от корня до любого таксона одинаково.
Вторым рассмотрим алгоритм Фитча-Марголиаша. Этот метод немного сложнее, чем UPGMA, но основан на том же подходе. Тем не менее, попытаемся отказаться от предположения UPGMA о молекулярных часах.
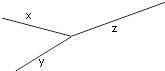
Прежде чем изложить алгоритм, сделаем несколько математических наблюдений. Во-первых, если попытаемся поместить 3 таксона на некорневое дерево, то будет только одна топология, которую необходимо учитывать. Кроме того, для 3 таксонов можем назначить желаемые длины ребер, чтобы точно соответствовать данным. Чтобы убедиться в этом, рассмотрим дерево на рисунке 5.9. Если есть некоторые данные о расстоянии
,
и
, то можно составить систему уравнений
,
,
.
Эти уравнения могут быть решены либо путем записи системы в виде матричного уравнения и нахождения обратной матрицы, либо путем подстановки формулы для одной переменной, полученной из одного уравнения, в другие. Любой способ гарантированно приведёт к следующему решению
,
,
.
Рисунок 5.9. Некорневое 3-таксонное дерево.
Будем называть эти формулы 3-точечными формулами для подгонки таксонов к дереву. К сожалению, с более чем 3 таксонами точная подгонка данных к дереву обычно невозможна. Однако алгоритм Фитча-Марголиаша (кратко называемый в таблицах как FM) использует случай 3 таксонов для обработки большего количества таксонов. Теперь объясним работу алгоритма на примере. Будем использовать данные о расстоянии, приведенные в таблице 5.4.
Таблица 5.4. Расстояния между таксонами
.31 1.01 .75 1.03
1.00 .69 .90
.61 .42
.37
Начинаем с выбора ближайшей пары таксонов для присоединения, как это делали в UPGMA. Глядя на таблицу расстояний,
и
являются первой парой, которая соединится. Чтобы соединить их, не помещая их на равное расстояние от общего предка, временно сводим задачу к случаю 3-таксонов, объединяя все остальные таксоны в группу. Таким образом, для имеющихся данных вводим группу
. Находим расстояние от каждого из
и
до группы, усредняя их расстояния до каждого члена группы. Таким образом, расстояние от
до
равно
, в то время как от
до
оно равно
. Это дает таблицу 5.5.