Со времен Дарвина человечество успело глубоко осознать опасности расовой концепции, столкнувшись с притеснениями и насилием, которые представители одной расы творили против других. Многие ученые, в том числе генетики, пытались преуменьшить степень биологического разнообразия внутри человеческого рода. Одной из самых влиятельных точек зрения на расу доныне остается позиция, обозначенная генетиком Ричардом Левонтином в 1972 г.
Левонтин измерил параметры белков, взятых у представителей разных рас (метода секвенирования ДНК еще не существовало), и вычислил стандартный показатель колебаний, известный как индекс фиксации по Райту, или FST. Смысл индекса в том, что у некоторого свойства, варьирующего в пределах популяции, подсчитывается доля вариаций, вызванная различием между двумя субпопуляциями. Таким образом, индекс Райта отражает ту долю вариаций выбранного признака, которую можно объяснить различиями между субпопуляциями.
Ричард Левонтин установил, что индекс Райта равняется 6,3 %, т. е. из всех различий между людьми, по крайней мере отражаемых 17 белками, которые Левонтин рассматривал, только 6,3 % представляют собой различия между расами, при этом еще 8,3 % – различия между этническими группами внутри рас. «На 85 % различия между отдельными людьми объясняются индивидуальным своеобразием человека внутри нации или племени», – заключил Левонтин{247}.
Это открытие вполне укладывается в теорию о том, что большая часть генетических вариаций в каждой расе совпадает с вариациями генофонда прародителей, откуда все и вышли. Но здесь также возникает вопрос: насколько велика степень различия между расами? Левонтин заявлял: межрасовые различия столь незначительны, что деление людей на расы с точки зрения генетики бессмысленно и не оправданно.
Такой интерпретации предпочитали держаться многие биологи, и позицию Левонтина приняли и даже довели до крайности крупнейшие общественно-научные ассоциации США. Согласно Американской ассоциации социологов, у расы нет биологической природы, поскольку это «социальный конструкт». Официальное коммюнике Ассоциации по вопросу рас гласит: «Признавая расовые категории как законный предмет эмпирического социологического исследования, важно сознавать всю опасность подыгрывания популярной концепции рас как биологического явления»{248}.
Американская антропологическая ассоциация также не признает, что между расами можно усмотреть биологические различия: «В США как ученые, так и широкая публика привыкли видеть в расах природное и объективное разделение человеческого рода на основе очевидных физических различий», – гласит ее декларация. Но поскольку физические особенности по всей Земле плавно варьируют и не связаны между собой, «эти обстоятельства делают все попытки провести четкие границы между биологическими популяциями субъективными и произвольными»{249}.
Однако сегодня людей можно объективно классифицировать по континенту происхождения, т. е. по расе, с помощью генетических маркеров, подобных тем, что использовала группа Фелдмана. И оценка Левонтином обнаруженных им различий как «незначимых» была столь же научным, сколь и политическим заявлением. Степень различия, исчисленная им в человеческой популяции, совпала с оценками других ученых, помещавших глобальное значение индекса Райта между 10 и 15 %. Сьюэлл Райт, один из трех основателей популяционной генетики и изобретатель индекса FST, отмечал: «…у других биологических видов столь заметно различные расы считались бы подвидами»{250}. Райт подчеркивал, что FST от 5 до 15 % в любой популяции живых организмов означает «умеренные» генетические различия, а диапазон от 15 до 25 % должен считаться «значительной» генетической дифференциацией{251}.
Однако существенную или не очень степень разнообразия отражает FST в 10–15 % – пожалуй, не так уж важно для ответа на вопрос, стоит ли разграничить человеческие расы. Дело в том, что замеры для индекса фиксации по Райту захватывают так называемые нейтральные вариации, а большинство расовых различий, скорее всего, программируются не ими.
Под нейтральными вариациями понимают мутации, никак не сказывающиеся на организме. Эволюцию подобного рода изменения не интересуют, и частотность таких аллелей в популяции колеблется стихийно от дрейфа генов. Общие вариации по большей части нейтральны, а большинство замеров FST, вероятнее всего, захватывают общие, т. е. нейтральные изменения.
Вместе с тем эволюция во все глаза следит за мутациями, изменяющими ген и программируемый им белок. Если изменения неблагоприятны, мутация безжалостно уничтожается, потому что ее носитель погибает или не может продолжить род. Если же модификация повышает репродуктивный успех индивида, мутация продвигается, так как становится все более частотной в популяции. Это и есть две стороны естественного отбора: отрицательный и положительный отбор. Биологи пока не знают, какие именно гены должны видоизмениться, чтобы один вид разделился на два подвида. Вместе с тем похоже, что аллели, задействованные в разделении человеческого рода на расы, скорее должны относиться не к нейтральным, а к закрепленным эволюцией.
Именно такую картину дают версии двух мозговых генов, сменившиеся в последние 40 000 лет их эволюции. Как мы упоминали в главе 4, аллель одного из этих генов, а именно микроцефалина, появившийся около 37 000 лет назад, сегодня широко распространен среди европеоидов и восточных азиатов, но гораздо реже встречается в Черной Африке. Индекс Райта для этого аллеля при сравнении Черной Африки и остального мира 48 %, или 0,48, «что указывает на большое разнообразие и существенно выше, чем среднегеномный показатель 0,12», – пишет первооткрыватель аллеля Брюс Лан из Университета Чикаго{252}.
Новая версия гена ASPM появилась около 6000 лет назад у европеоидов, среди которых носители этого аллеля составляют 44 %. В Восточной Азии он менее распространен, а в Черной Африке крайне редок. Индекс FST этого аллеля у европеоидов и неевропеоидов расходится на 0,29{253}. Несомненно, существуют аллели других мозговых генов, еще не известные науке, которые широко распространены среди азиатов и африканцев и редки у европейцев.
Например, различия в цвете кожи у людей по большей части межпопуляционные, а не внутрипопуляционные. По данным анализа, проведенного Джоном Релетфордом из Университета штата Нью-Йорк в Онеонте, FST по цвету кожи составляет 88 %{254}. Цвет кожи сильно зависит от расы и от географической широты, но очевидно не диктуется нейтральными генами. Генетики Генри Харпендинг и Алан Роджерс считают, что «с другими заметными признаками, отмечаемыми большинством наблюдателей, дело, скорее, обстоит так же, как с цветом кожи, чем как с нейтральными модуляциями». Иначе говоря, большинство физических характеристик, по которым публика определяет расу человека, скорее всего, закреплены отбором, что и было бы закономерно, если главной силой, задававшей своеобразие разных человеческих популяций, был половой отбор{255}.
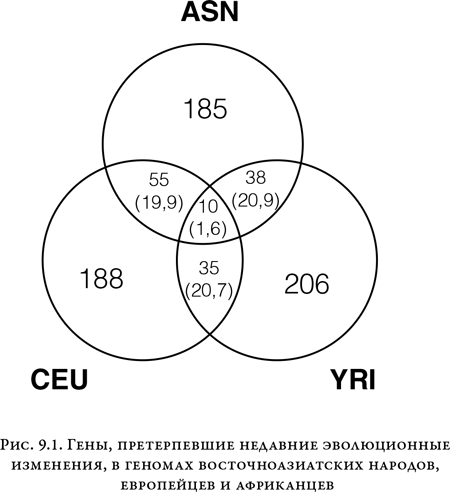
Именно выбранные эволюцией гены, а не множество нейтральных, скорее всего, ответственны за межрасовые различия. Убедительные свидетельства в пользу этой гипотезы обнаружил пангеномный анализ, осуществленный Джонатаном Притчардом из Университета Чикаго. Разрабатывая тест на выявление генов, недавно подвергшихся селективному давлению, Притчард нашел примерно по 200 таких генов у африканцев, азиатов и европейцев. Лишь совсем незначительная часть из этих наборов у трех рас совпадает, чего и следовало бы ждать, если людям на разных континентах приходилось приспосабливаться к эволюционному давлению независимо друг от друга{256}.