Вскоре Уилсон и его сотрудники сконцентрировались на ДНК определенного типа. Большая часть ДНК хранится в ядрах наших клеток, но небольшая доля находится в митохондриях — одном из типов органелл, расположенных в цитоплазме клетки между ядром и мембраной и отвечающих за клеточное функционирование. В митохондриях содержится гораздо меньше ДНК, чем в ядре, но клетка имеет несколько митохондрий, а значит, одна и так же молекула ДНК дублируется в ней несколько раз. Так называемая мтДНК имеет огромный потенциал для молекулярной систематики. Большая часть ядерной ДНК перемешивается после слияния яйцеклетки и сперматозоида, поэтому проследить ее между поколениями невозможно. Митохондриальная же ДНК дает четкое представление о наследственности, потому что она передается без всяких изменений по материнской линии (сперматозоиды отца, по сути, представляют собой всего лишь ядра клеток). Соответственно, любые различия в мтДНК между двумя особями, когда-то имевшими общего предка, объясняются исключительно мутациями (то есть спонтанными изменениями, постоянно возникающими в четырех возможных типах нуклеотидов).
В 1987 году Уилсон и его студенты Ребекка Кэнн и Марк Стоункинг исследовали мтДНК 147 человек из разных регионов мира. Предположив, что скорость изменчивости должна быть постоянной, и приняв во внимание количество различий между образцами, они рассчитали, что наш вид возник около 200,000 лет назад. Более того, полученное ими «дерево мтДНК» указывало на происхождение Homo sapiens с африканского континента. Это подтверждалось и тем фактом, что потомки жителей Африки имеют наибольшее разнообразие мтДНК, то есть люди живут и развиваются здесь дольше, чем где бы то ни было еще. Относительно точной даты возникновения Homo sapiens в молекулярной систематике ведутся жаркие споры, но два факта несомненны. Во-первых, наш вид появился относительно недавно, а во-вторых, через какое-то время после возникновения он прошел через «бутылочное горлышко», то есть существенное сокращение популяции, после чего покинул африканский континент.
Меня, естественно, очень привлекали эти открытия, и не только потому, что они подтверждали мои собственные размышления о палеонтологической летописи. Молекулярный подход Уилсона и его коллег указывал на то, что, даже если наш вид еще не достиг конца своего развития, оно по крайней мере имело четкое начало. Невозможно было заниматься молекулярной систематикой, не признавая, что виды действительно существуют, а такое признание давало возможность для более тонкого анализа палеонтологической летописи человечества. Но, чтобы научное сообщество могло это осознать, требовалось время. Должен признаться, в тот момент меня больше всего радовало, что неандертальцев с их уникальными характеристиками наконец-то признают отдельным видом, а не просто вымершей разновидностью Homo sapiens.
Я полагал, что неандертальцы существенно отличались от нас не только с анатомической, но и с поведенческой точки зрения, что подтверждалось археологическими находками. Мне казалось, что, отрицая уникальность этих далеких родственников человека, палеоантропологи отказывались от возможности понять их в их собственном контексте, как отдельный вид, которым они почти наверняка и являлись. В конце концов, мне надоело ждать, пока эксперты по неандертальцам разберутся с этой проблемой, и в 1986 году я написал работу, посвященную трудностям классификации окаменелостей по их морфологии. В ней подчеркивалось, что видообразование как процесс, результатом которого является возникновение новых биологических единиц, способных к размножению, очевидно, не пассивное последствие морфологических изменений. Одни виды накапливают множество таких изменений, но не разделяются на несколько разных частей, а другие, наоборот, едва различаются на вид, но при этом совершенно несовместимы в репродуктивном плане. Эти явные различия между морфологией и статусом вида не играют большой роли, когда у вас есть возможность наблюдать за взаимодействием (или его отсутствием) живых существ. Но вот если особи, которых вы анализируете, живут вдали друг от друга, или же если у вас имеются только их окаменелые кости, такие различия становятся крайне важным фактором.
Практический опыт (в том числе и моя работа с лемурами) показывал, что зачастую родственные виды приматов не имеют значительных различий в морфологии костей и зубов. Таким образом, если вы обнаруживаете в одном раскопе две явно различающиеся окаменелости твердых тканей, высока вероятность, что перед вами останки двух разных видов, разумеется, если вы сможете исключить половой диморфизм. Более того, так как различия между костями родственных видов обычно незначительны, палеоантропологи легко могут недооценить количество видов, представленных тем или иным собранием окаменелостей. Разумеется, мои рассуждения вводили некоторый элемент неуверенности, но даже он был предпочтительнее, чем излишнее упрощение эволюционных паттернов вместо их классификации. С другой стороны, если я был прав, то все попытки выделить на основании окаменелостей подвиды, то есть четко выраженные региональные варианты, возникающие внутри видов и быстро исчезающие в них же, были обречены на провал. Подвиды — это движущая сила эволюционных инноваций, основа для формирования новых видов. Но до сих пор, пока процесс видообразования не наделит их исторической индивидуальностью, они остаются эфемерными.
Практические трудности, связанные с распознаванием близко-родственных видов в палеонтологической летописи, никуда не делись с 1986 года, когда моя работа была опубликована. Я приложил все усилия, но так и не сумел найти способ, как их устранить. Тем не менее положительным результатом моей публикации стало то, что в последние десятилетия палеоантропологи все реже и реже употребляют термин «стадия». Стадия — это особая палеоантропологическая концепция, которую я однажды назвал самой ужасной «уткой», выдуманной моими коллегами. Стадией называют неопределенную группу окаменелостей, имеющих примерное сходство (например, по объему мозга). Такая группировка, разумеется, является продуктом человеческого сознания, а не природы. Стадии не существуют сами по себе, это всего лишь эвристические инструменты, которые тем не менее иногда путают с реальностью. Сам по себе этот термин не так уж плох, но если палеоантрополог мыслит «стадиями», он забывает о естественной классификации, созданной самой природой. Если кто-то говорит, что эволюция гоминидов прошла через «стадию Homo erectus», то подразумевается, что в ней отсутствуют такие понятия, как предки и наследование. Вместо этого всю работу выполнил естественный отбор, и дальнейшие исследования не имеют смысла. Придерживаясь идеи «стадии», мы проигнорируем тот факт, что за последние пару миллионов лет рост объема мозга в роду Homo происходил независимо в рамках трех разных линий развития: у Homo erectus в Восточной Азии, у предков неандертальцев в Европе и у предков Homo sapiens в Африке. Это очень важно, так как если рост мозга является свойством всего рода Homo в целом, а не конкретно нашего вида, то и корни этого явления нужно искать совершенно в другом месте.
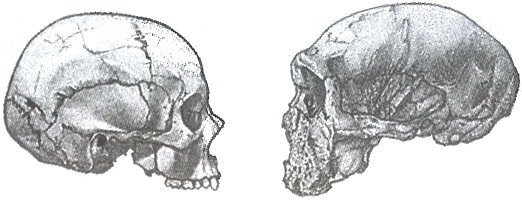
В 1986 году моей главной задачей было показать, что Homo sapiens представляет собой во многом уникальный вид и что лишь некоторые окаменелости (имеющие сравнительно молодой возраст) вписываются в его морфологическую структуру. В палеонтологической летописи плейстоцена отсутствует и большая круглая черепная коробка, и достаточно маленькое лицо в ее нижней части, характерные для современного человека. Таким же уникальным является и Homo neanderthalensis с его длинным списком характеристик, не похожих ни на чьи другие (близкие родственники неандертальцев из Сима-де-лос-Уэсос к тому моменту еще не были обнаружены).
Различия в строении черепа между современным Homo sapiens (Эль-Хеса, Египет, слева) и Homo neanderthalensis (Гротта-Гуаттари, Монте-Сирсео, Италия). В масштабе. Рисунки Дона Макгрэнэгана (слева) и Дианы Саллес (справа)