Новое видение
После четырех лет в Йеле, в 1971 году, мне посчастливилось получить работу младшего куратора в Американском музее естественной истории в Нью-Йорке. К моменту выпуска из университета я уже знал достаточно, чтобы понимать, что меня так и не научили правильно читать те окаменелости, с которыми я работал. Я уже с большей уверенностью делал выводы, но все еще не мог точно определить, чем я занимаюсь. Некоторое время это вообще казалось мне не таким уж важным. Я написал диссертацию по семейству вымерших лемуров, и одним из преимуществ этой темы оказалась поездка на Мадагаскар для изучения коллекции окаменелостей в Малагасийской академии. Во время своего пребывания там я воспользовался возможностью прогуляться по лесам и понаблюдать за обитающими там лемурами. Я тут же влюбился в этих очаровательных созданий, и мои планы на будущее очень скоро начали вращаться вокруг живых лемуров, а не их вымерших предков. Я хотел узнать больше о жизни этих далеких предков человечества, которые в то время были практически не изучены, а также понять причины их огромного разнообразия. Я хотел выяснить, какие именно виды лемуров обитали на этом острове, как они жили и почему находились именно здесь. Чтобы ответить на все эти вопросы, мне пришлось вступить в совершенно новую область исследований, сильно отличавшуюся от мертвого мира окаменелостей, в котором я жил раньше.
К несчастью или нет, моя карьера полевого исследователя лемуров длилась недолго. В 1972 году проколониальное правительство Мадагаскара было свергнуто, и к середине 1974 года остров погрузился в хаос. К концу года меня депортировали, и так я впервые оказался на Коморских островах, о которых говорилось в самом начале книги. К середине 1975 года я вернулся в Нью-Йорк к своим окаменелым останкам гоминидов, которые я забросил несколько лет назад. Это был удачный момент, так как в начале 1970-х годов в Американском музее естественной истории тоже назревала революция, на сей раз в систематике.
Еще в 1950 году немецкий энтомолог Вилли Хенниг опубликовал новаторскую работу о процедуре систематизации, в которой описал очень четкий подход к биологической классификации и реконструкции эволюционных взаимоотношений. Вездесущий Эрнст Майр чуть позже назвал эту систему кладистикой (от древнегреческого корня, означающего «ветвь», так как для обозначения взаимоотношений в ней использовались разветвленные диаграммы). Но лишь после 1966 года, когда книга Хеннига «Филогенетическая систематика» была переведена на английский, о кладистике стало известно за пределами Германии. Ее популярности в англоязычном мире поспособствовали два ихтиолога из Американского музея естественной истории, Донн Роузен и Гарет Нельсон, неутомимо отстаивавшие новую точку зрения во времена, когда большинство систематиков придерживались той или иной версии «общего морфологического паттерна» Уилфрида Ле Грос Кларка. Традиционно взаимоотношения между организмами оценивались на основании интуитивного восприятия общего сходства, и в случае возникновения споров между двумя авторитетными систематиками их точки зрения было очень сложно сравнить между собой. Выбор между двумя альтернативами сводился к тому, чье заявление казалось слушателям более убедительным, поэтому умение говорить ценилось выше, чем стройные рассуждения.
Несмотря на то что многие традиционные систематики тут же заявили, что уже давно применяли подход Хеннига, на самом деле он оказался совершенно уникальным. Хенниг создал четкую систему, позволявшую (по крайней мере теоретически) сравнивать конкурирующие гипотезы родства. Главная идея Хеннига состояла в том, что не все морфологические сходства созданы равными или по крайней мере их важность меняется со временем. Разумеется, систематики уже давно знали, что не все морфологические характеристики можно в равной степени использовать для разработки теорий родства. Они различали синапоморфические свойства, унаследованные от одного предка и могущие служить основанием для установления родства, и конвергенцию, то есть независимое (и малоинформативное с точки зрения систематики) приобретение внешне сходных признаков. Классическим примером конвергенции являются крылья птиц и летучих мышей. Однако Хенниг пошел еще дальше и указал, что даже характеристики, полученные от одного предка, могут иметь разную важность для систематизации. После того как систематик использует какую-то черту для различения предка и его потомков, такая черта теряет свою значимость при определении взаимоотношений между потомками. В конце концов, она есть у каждого из них. Соответственно, для выявления родства между членами группы нужно искать иные общие черты или альтернативные варианты одной характеристики.
К примеру, наличие у животного четырех лап указывает на его принадлежность к группе четвероногих, к которой относятся все наземные позвоночные. Но это свойство никак не указывает на то, какие группы четвероногих имеют более близкое родство друг с другом. Соответственно, для анализа взаимоотношений внутри группы эту общую характеристику можно опустить. Иными словами, сходные черты (будь то морфологические или молекулярные), унаследованные от общего предка, могут быть примитивными или производными в зависимости от того, насколько близким был предок. Любая унаследованная от предка морфологическая характеристика (например, наличие четырех лап) может быть производным свойством предка, объединяющим широкую группу потомков и отличающим ее от других. Однако внутри этой группы такая черта будет считаться примитивной, так как она будет иметься у всех потомков и ее невозможно будет использовать для анализа внутригрупповых отношений. Суть всей идеи заключалась в том, что эволюционные взаимоотношения можно установить только на основании общих производных характеристик. Придавая одинаковую значимость всем сходным чертам, вы рискуете совершить ошибку.
Но как же определить, какие черты группы являются примитивными, а какие — производными? Для этого существует несколько способов. Во-первых, примитивные характеристики будут широко распространены в исследуемой вами группе, в то время как производные будут встречаться гораздо реже. Более того, если какая-то черта встречается у близких родственников исследуемой вами группы, это означает, что она также является примитивной и не может использоваться для выделения подгрупп. Очень часто помогает и анализ развития той или иной черты. Тот факт, что и у рыб, и у человека на ранних стадиях эмбрионального развития присутствуют жаберные щели, делает это свойство производным для всех современных позвоночных (несмотря на то что у их общего предка оно являлось производным). Очень часто палеонтологи считают, что характеристики, возникшие на ранних этапах развития группы, являются примитивными, а более поздние — производными. Однако такая логика может оказаться достаточно рискованной.
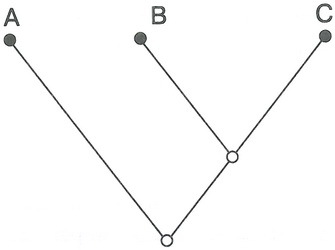
Пример простой кладограммы. Черные кружки означают три существующих таксона, к примеру вида. Линии указывают на реконструированные генеалогические отношения между ними. Пустые кружки означают гипотетическое родство, основанное на наличии у видов В и С характеристик, отсутствующих у А. Иными словами, пустые кружки не соответствуют существующим в природе таксонам
Для создания схемы взаимоотношений, или кладограммы, нужно собрать воедино все интересующие виды, как живые, так и вымершие. Используя все характеристики таких видов, вы выявляете те, которые присутствовали у общего предка всей группы и предков подгрупп внутри нее. Сами по себе эти предки могут быть неизвестными, и вы даже можете не располагать их окаменелыми останками, но их характерные черты должны быть ярко выражены. В результате должна получиться четкая схема родства всех организмов в группе с общим предком. Кладограмма может быть впоследствии признана неверной или изменена путем добавления новых морфологических характеристик, таксонов или ваших собственных рассуждений. Кроме того, кладограмму можно применять для сравнения с другими сходными или конкурирующими гипотезами на основании анализа характеристик, использованныхв каждой из них.