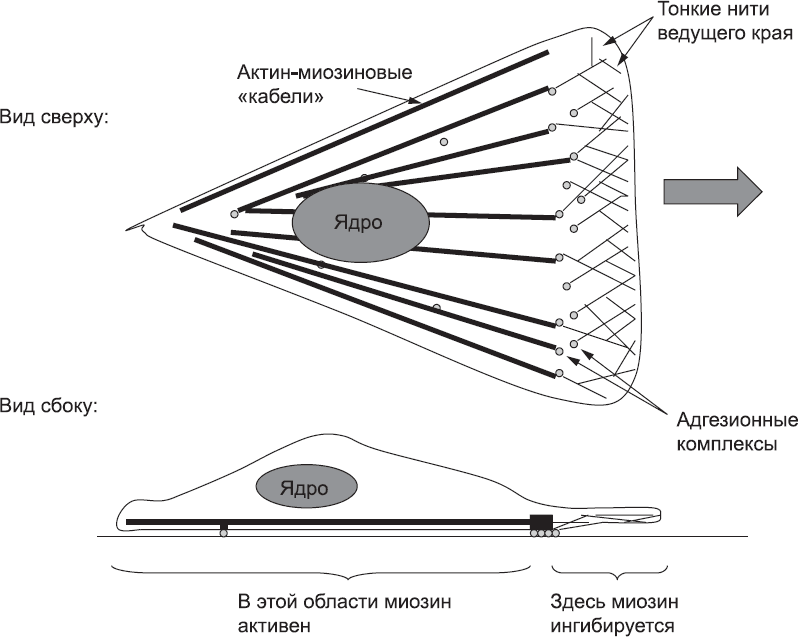
Таким образом, небольшой набор компонентов – актин и несколько белков, регулирующих его поведение, – может использовать для самоорганизации простую асимметрию клеточной среды (степень удаленности конкретной части клетки от мембраны) и создавать «двигатель», обеспечивающий перемещение клетки. Все эти белки, конечно, кодируются генами в ядре, но, помимо производства белков, ядро и гены не принимают никакого участия в создании «двигателя».
Рис. 35. Схема расположения тонких микрофиламентов у переднего края клетки, адгезионных комплексов в задней его части и актин-миозиновых «кабелей», идущих в заднем направлении
Это было прекрасно продемонстрировано в экспериментах с клетками рыб: небольшие фрагменты клеток, лишенных ядра (а значит и генов), были вполне способны к перемещению. Обеспечивает эту способность набор белков – именно набор, а не какой-то конкретный белок.
Итак, мы ответили – по крайней мере, в общих чертах – на первую часть вопроса о клеточных миграциях. Мы поняли, за счет чего клетки вообще способны к движению. Теперь нужно понять, как они могут перемещаться в определенном направлении. Клетки эмбриона используют два подхода к решению этой проблемы (либо по отдельности, либо, что бывает чаще, вместе). Один заключается в предоставлении клеткам выбора между поверхностями с разными адгезионными свойствами, а другой – в работе молекул, которые усиливают активацию белков, инициирующих полимеризацию актина, в передней части клетки.
Организацию направленного перемещения клеток за счет дифференциальной адгезии проще всего рассмотреть на примере ряда экспериментов последних десятилетий ХХ века. В лабораторных условиях клетки можно выращивать на самых разных поверхностях – от пластика до сложных биологических молекул. Адгезионные свойства этих поверхностей можно расположить в порядке возрастания просто на основании того, насколько легко или трудно смыть клетки с той или иной поверхности струей воды. Когда клетки помещают на поверхность с «мозаичной» текстурой (некоторые части обладают большей адгезией, чем другие), они беспорядочно перемещаются, пока не достигнут границы. Там клетки могут выбрать, куда им двигаться, и они выбирают поверхность с более сильной адгезией. Этому есть простое объяснение. Рассмотрим клетку, которая расположена на границе скользкой и липкой поверхностей (рис. 36). По определению, та часть, которая находится на более липкой поверхности, будет формировать больше адгезионных контактов, чем другая часть. Сеть тонких филаментов в области переднего края будет хорошо «заякорена», и лидирующий край клетки, отталкиваясь от этого якоря, будет быстро продвигаться вперед. На скользкой поверхности такого «якоря» нет, поэтому лидирующий конец будет расходовать силы на движения в обратном направлении.
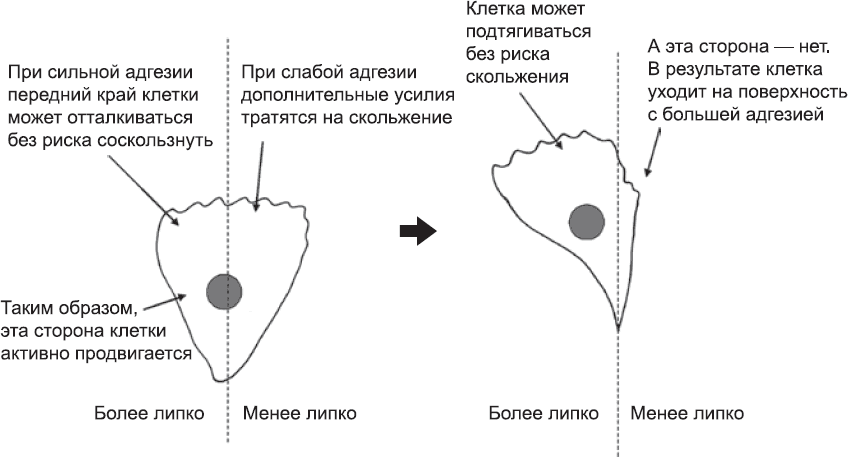
Рис. 36. Оказавшись на границе между двумя поверхностями, одна из которых благоприятствует формированию адгезионных комплексов, а другая – нет, клетка переместится на поверхность с более высокой адгезией
Актин-миозиновые «кабели», которые подтягивают основную часть тела клетки вперед, тоже будут лучше крепиться на липком субстрате, что облегчит движение тела клетки в этом направлении. Поэтому и лидирующий край клетки, и ее тело будут заведомо лучше двигаться по липкой поверхности, а значит, туда-то клетка и направится.
В течение нескольких лет ученые сомневались в том, что направленное перемещение клетки за счет адгезии основано на чисто механическом взаимодействии. Высказывались предположения, что сигналы от адгезионных комплексов стимулируют белки, инициирующие полимеризацию актина, то есть направленное перемещение происходит за счет химических процессов. Сомнения развеял один изящный эксперимент,[121] в ходе которого исследователи поместили крошечный стеклянный шарик, покрытый молекулами с хорошей адгезией, на верхнюю поверхность лидирующего края клетки. Шарик прилип к адгезионным комплексам, и сеть актиновых филаментов лидирующего края пыталась использовать его в качестве «якоря» для отталкивания. Шарик не был ни к чему прикреплен, поэтому он просто откатывался назад, а лидирующий край клетки вперед не продвигался. На направление движения клетки это не влияло. Однако когда авторы эксперимента зафиксировали шарик при помощи тончайшей стеклянной нити, лидирующий край клетки начал быстро продвигаться вперед, и клетка стала двигаться в направлении шарика. Химические сигналы были одинаковы в обоих случаях, механическая сила появлялась только тогда, когда шарик был закреплен. Этот эксперимент убедительно показал, что направленное движение клеток в принципе может происходить только под действием механических сил в отсутствие каких-либо химических различий.
Тем не менее, хотя в ряде случаев направленное движение клеток действительно происходит чисто механическим путем, в других случаях химическая сигнализация играет очень важную роль. Некоторые молекулы на поверхности не используются для адгезии, а распознаются специальными рецепторами на клеточной мембране, которые активируют сигнальные пути внутри клетки. Некоторые рецепторы и сигнальные пути локально стимулируют активность белков, инициирующих полимеризацию актина, а значит, способствуют локальному продвижению лидирующего края клетки. Другие действуют прямо противоположным образом, подавляя продвижение лидирующего края; в этом случае клетки избегают подобных поверхностей.
Эксперименты со стеклянными шариками и мозаичными поверхностями – это любопытно, но как связана склонность клеток выбирать более липкую поверхность с клеточными миграциями в сложной внутренней среде эмбриона? Если, например, клетки просто выбирают поверхность с более сильной адгезией, то как могут разные клетки одновременно следовать разными маршрутами (которые к тому же иногда пересекаются)? Ответ на этот вопрос отчасти заключается в том, что существует множество разных типов адгезионных комплексов, и каждый из них сильнее всего связывается с определенным поверхностным белком. Например, адгезионный комплекс α6β1-интегрин связывается с поверхностным белком ламинином, а аналогичный комплекс α5β1-интегрин связывается с другим поверхностным белком – фибронектином. Разные типы клеток несут специфические комбинации адгезионных комплексов. Клетки с адгезионным комплексом α6β1-интегрином будут «считать» ламинин поверхностью с сильной адгезией, а клетки с α5β1-интегрин – не будут. Это означает, что разные типы клеток будут выбирать разные направления движения, даже если они движутся по одним и тем же поверхностям.
Неподвижные клетки развивающихся тканей эмбриона синтезируют поверхностные молекулы, такие как ламинин и фибронектин. При этом разные типы клеток секретируют поверхностные молекулы в разных сочетаниях. Это означает, что ползущая клетка воспринимает разные ткани эмбриона как местность с разнообразным и мозаичным рельефом: одни поверхности более липкие, другие – менее липкие, некоторые также несут сигнальные молекулы, привлекательные или непривлекательные для клетки. Разные типы ползущих клеток «видят» этот ландшафт по-разному в зависимости от того, какие адгезионные комплексы и рецепторы они несут, и поверхность, исключительно привлекательная для одного типа клеток, может оказаться абсолютно непривлекательной для другого типа клеток. Это еще один пример того, что смысл знака зависит от интерпретации. Полной информации об означаемом знак сам по себе не несет. Таким образом, каждый тип клеток может продвигаться по эмбриону своим путем.
Мы постоянно узнаем что-то новое о путях миграций клеток по эмбриону, однако того, что мы уже знаем, достаточно для того, чтобы проиллюстрировать только что изложенные общие принципы конкретными примерами. Каждый из описанных случаев относится к популяции клеток, которая отделяется от самой дорсальной части нервной трубки и мигрирует, распадаясь на потоки неплотно соединенных клеток, к разным частям организма. Эти клетки названы по месту их происхождения «клетки нервного гребня», и разные популяции этих клеток имеют разную судьбу. Они дают начало, во-первых, сенсорным клеткам, отвечающим за передачу спинному и головному мозгу тепловых, болевых и тактильных ощущений, а также информации о местоположении, во-вторых, вегетативной нервной системе, отвечающей за неподконтрольные сознанию функции органов, например частоту сердечных сокращений. В-третьих, клетки нервного гребня формируют основную часть надпочечной железы, которая выделяет гормоны (например, адреналин). В-четвертых, они дают начало пигментным клеткам, защищающим кожу от ультрафиолетового излучения. Разные судьбы клеток нервного гребня связаны с разными путями миграции.