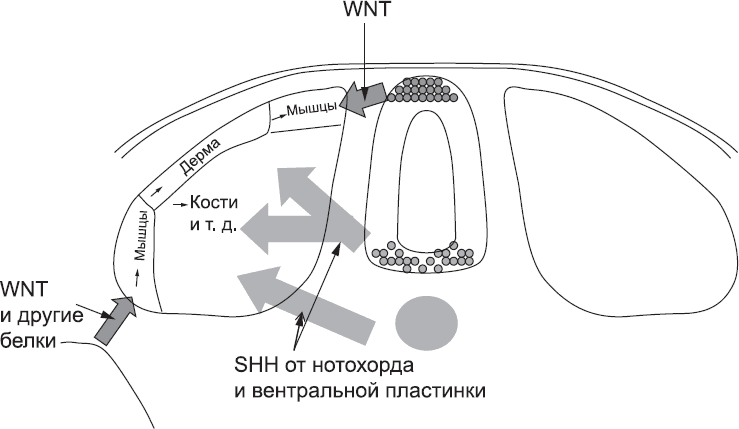
Когда клетки сомита получают сигналы WNT из дорсальной части нервной трубки, они начинают производить белки, необходимые для формирования мышц. Белок WNT распространяется лишь на довольно небольшое расстояние, поэтому сигнал для активации развития мышц получает только та часть сомита, которая находится ближе к нервной трубке. WNT, поступающий из другого источника, запускает образование второго центра развития мышц на наружном, нижнем крае сомита (рис. 32). В дальнейшем из клеток этих зон образуются разные мышцы: зона около нервной трубки даст начало мышцам спины, а другая зона – мышцам стенки тела.[112] Зона между ними, расположенная вдали от обоих источников WNT, станет соединительной тканью кожи (дермой).
Если бы на сомит действовали только те три типа сигналов, о которых мы только что говорили, он стал бы похож на сэндвич, в котором зона, дающая начало коже, залегает между двумя зонами, образующими мышцы.
Рис. 32. Разметка областей сомита, которые будут дифференцироваться в разных направлениях, происходит за счет сигналов от соседних тканей
Однако сомит также чувствителен к сигналам, поступающим от нотохорда и вентральной пластинки нервной трубки. В участках сомита, расположенных ближе всего к этим источникам сигналов, концентрация соответствующих сигнальных белков достаточно высока. Они блокируют сигналы, стимулирующие образование мышц и кожи, и «сообщают» клеткам, что они должны образовать соединительную ткань и кости[113],[114] (рис. 32).
Итак, обобщая сказанное в этой главе, мы видим, что окружающие ткани подают сигналы, за счет которых происходит дифференциация нервной трубки, а клетки нервной трубки реагируют на эти сигналы, в том числе и тем, что подают сигналы, под действием которых происходит разметка окружающих тканей и они приступают к дифференцировке. Таким образом, тонкая детальная разметка строения эмбриона, который еще недавно имел довольно однородное строение, является результатом многочисленных «обменов репликами» на клеточном уровне.
Ткани, о которых шла речь в этой главе, не уникальны. Те же самые общие принципы можно было бы продемонстрировать на примере любой другой группы соседних тканей. Во всем эмбрионе смежные участки тканей используют друг друга как источник сигналов для выделения внутри себя областей, клетки которых будут подвергаться дифференцировке, характер которой зависит от их расстояния до источника сигнала. Непосредственным результатом этого процесса является образование новых границ между только что дифференцировавшимися типами клеток в пределах исходно гомогенного участка ткани. Если эти типы клеток секретируют разные сигнальные белки, тот же прием можно использовать еще раз для создания еще большего количества отличающихся друг от друга областей. Это высокоэффективный механизм пространственной дифференцировки, и неудивительно, что примерно пятая часть генов человека нужна для производства белков, вовлеченных тем или иным образом в процесс клеточной сигнализации.
Клеточная коммуникация не только позволяет тканям дифференцировать внутри себя практически неограниченное число областей. Она также служит отличным механизмом борьбы с ошибками в процессе развития. Представьте себе эмбрион, в котором дифференциация участков тканей не зависела бы от положения окружающих тканей, а происходила бы каким-то иным путем, например, клетки действительно следовали бы пошаговым инструкциям к какому-то чертежу. Малейшие неточности в положении конкретных клеток приводили бы к постепенному накоплению ошибок; когда пришло бы время тканям собираться вместе, типы клетки, которые должны были бы соседствовать, оказались бы в разных местах, и развитие остановилось бы. В принципе, таким образом можно построить очень маленький и простой организм – при малом числе типов тканей и жизненно важных связей между ними ошибки просто не успеют накопиться. Однако создать «по инструкциям» организм, состоящий из сотен разных типов клеток, которые должны тонко взаимодействовать друг с другом, нельзя. Напротив, если ткани в системе полагаются на сигналы друг друга, дифференциация клеток в каждой из них автоматически происходит на правильном расстоянии от ткани, сигналы которой управляют этим процессом, даже если, скажем, эта «управляющая» ткань находится немного не там, где надо. Таким образом, организация системы подстраивается к меняющимся обстоятельствам, и ошибки не накапливаются, а корректируются на каждом этапе. Поэтому эмбрион способен справиться практически со всеми ошибками, за исключением серьезных аномалий, – постоянное «общение» клеток позволяет регулировать развитие в соответствии с реальной ситуацией, а не с абстрактным планом.
Использование общения на языке белков для регуляции дифференцировки тканей имеет одно интересное следствие для развития животных. Расстояние, на которое может распространяться белок в рабочих концентрациях, ограничено биофизическими и биохимическими законами и составляет для большинства белков около одной двадцатой доли миллиметра (50 мкм). Это означает, что длина группы клеток, использующих этот метод для нанесения разметки, предшествующей клеточной дифференцировке, как правило, составляет примерно 1/20 мм. Это относится как к расстоянию между дорсальной и вентральной частями нервной трубки, так и, например, к расстоянию между развивающимися корнями зубов или волосками. Из этого следуют две вещи. Во-первых, невозможно нанести разметку на весь эмбрион сразу. Сначала, пока эмбрион еще достаточно мал, нужно нанести грубую разметку, а потом, когда уже намеченные части увеличатся в ходе роста эмбриона, можно приступать и к их детализации. Это одна из причин того, почему человеческое развитие не может идти по пути создания крошечного, но полноценного ребенка, который затем только увеличивался бы в размерах. Последовательные фазы детализации должны быть распределены по этапам роста. Сначала обособляются голова и туловище, потом, когда голова вырастает, намечается, например, челюсть, затем, когда увеличивается челюсть, намечается положение зубов и так далее.
Второе следствие заключается в том, что нанесение любой разметки, например выделение зон нервной трубки, может произойти только на определенном этапе развития, когда размер эмбриона находится в довольно строгих границах. В частности, это означает, что на этом этапе развития зародыши землеройки, человека и синего кита должны быть практически одного размера. Эволюционное сходство животных – скажем, лошадей, китов и летучих мышей, которые, будучи очень разными, относятся к одному и тому же классу, – гораздо отчетливее видно на эмбрионах, чем на взрослых особях.
Глава 8
Внутренние путешествия
Когда дорога вся в цветах, уже не спрашивают, куда она ведет.
Анатоль Франс
Те клетки, которые сформируют лицо взрослого человека, располагаются в задней части головного отдела эмбриона. Те клетки, которые станут чувствительными нервами и пигментными клетками кожи, сначала находятся в районе позвоночника. А те клетки, которые стали вашими сперматозоидами или яйцеклетками, вообще какое-то время лежали за пределами вашего тела. Эти и многие другие подобные факты подчеркивают зависимость нашего развития от способности клеток раннего эмбриона перемещаться из одного места в другое. Миграции на клеточном уровне не менее удивительны, чем великие путешествия птиц и рыб, издавна восхищавшие зоологов, а возможно, и более, если учесть, что клетки, в отличие от птиц, перемещаются в постоянно меняющейся среде и делают это с исключительной точностью, хотя у них нет ни глаз, ни мозга, ни возможности учиться у родителей.