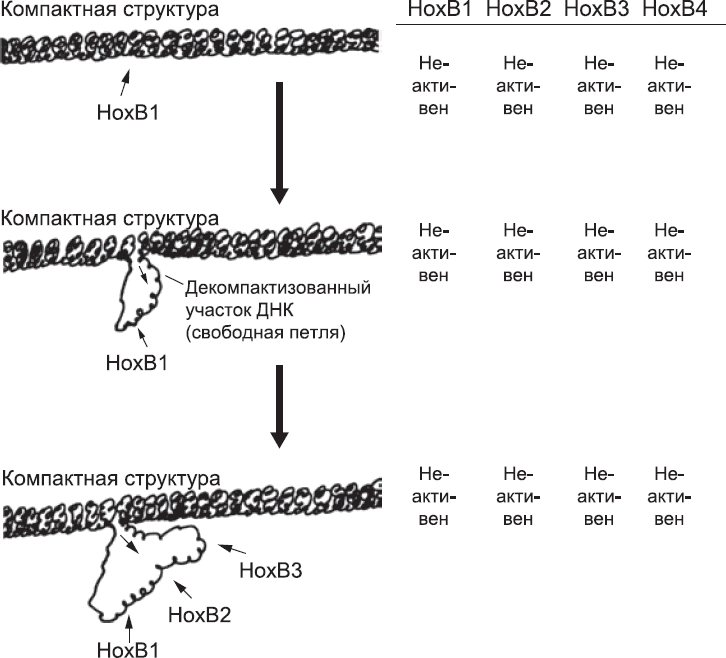
Перед активацией hoxb-генов кластер в компактном состоянии. Из всех генов hoxb-кластера самый правый ген (hoxb13) «похоронен» в глубине компактного клубка. hoxb1 наиболее доступен, будучи в начале компактной зоны. Экспрессия генов hoxb зависит от освобождения из этой «компактной тюрьмы». Это может случиться, если специфичные последовательности ДНК-связывающих белков присоединятся к ДНК с наиболее доступного конца и «перепрограммируют» упаковочные белки, так чтобы они позволили ДНК принять более свободную, петлеобразную форму. Эти ДНК-связывающие белки активируются ретиноевой кислотой. Эффект ретиноевой кислоты на hoxb-кластер настолько мощный, что его можно разглядеть даже в мышиных СКЭ, выращенных в простой чашке Петри.[95] Если постепенно добавить туда небольшое количество ретиноевой кислоты, «спрессованный» hoxb-кластер начинает ослабевать с конца. Так что hoxb1, hoxb2, hoxb3 и так далее будут доступны для транскрипции (рис. 29). Все это требует времени, что не удивительно, учитывая, что в ДНК hoxb-кластера около 150 000 оснований. Освобождение HOXb гена – постепенный процесс. Необходимо осуществлять контроль над порядком их «укладки» в кластере. Вероятно, то же самое верно[96] для hoxa-, с – и d-кластеров мыши и, по аналогии, для всех кластеров HOX человека.
Получение петли из hoxb-кластера также наблюдается в эмбрионе мыши. Скорость «разматывания» и волны активации генов совпадают с той скоростью, с которой гаструляция распространяется по направлению к «хвосту».[97] Таким образом, можно смело надеяться на то, что описанный выше механизм верен, хотя мы уже знаем, что его сопровождает огромное количество дополнительных сложностей. Необходимо убедиться, что клетки «запоминают» те гены для экспрессии, и это должно быть согласовано с соседними клетками. При этом отключаются гены слева, а те, что справа, подвергаются экспрессии. Тщательная систематизация всего этого процесса может стать делом жизни многих исследователей.
Человеческие HOX-гены (как и мышиные hox-гены) очень важны, поскольку существует связь между набором HOX-генов, экспрессированных сомитом, и типом будущего позвонка.[98] Например, третий, четвертый и пятый позвонки шейного отдела очень похожи: образовавшие их клетки экспрессируют тот же набор генов. Следующие два позвонка от них отличаются, и их клетки уже экспрессируют другие наборы HOX-генов.
Рис. 29. Постепенное изменение структуры HOX-кластера, от компактной до более рыхлой, высвобождает большее количество генов для транскрипции в том порядке, в котором они расположены в кластере
У различных видов животных число позвонков варьируется. У кур, например, четырнадцать шейных позвонков, а у мышей, как и у нас, – семь. В обоих случаях тем не менее начало экспрессии генов типа HOX6 (Hoxa6, Hoxb6 и т. д.) знаменует конец сомитов, создающих шейные позвонки, и начало тех, которые образуют позвонки грудного отдела. Поэтому гены HOX коррелирует с типом позвонков, а не просто с их числом. Аналогично начало экспрессии генов типа HOX10 означает конец сомитов, идущих на грудные позвонки, и начало тех, которые создают позвонки поясничного отдела. Гены типа HOX11 отвечают за переход от поясничного к крестцовому позвонку. Все это подтверждает, что тип HOX-генов может определять вид позвонков, который создадут сомиты.
Эта идея была проверена экспериментально в ходе генной инженерии эмбрионов мыши. Из них убирали один или несколько Hox-генов и наблюдали последствия этой операции для развития. В шейном отделе нормальной мыши должен быть один первый позвонок («атлант») и один второй («ось»). У каждого из них своя специфика, они позволяют по-разному осуществлять кивки и вращения головы. У третьего, четвертого и пятого позвонков форма, характерная для позвонков шейного отдела. При удалении гена Hoxa4 третий, неспециализированный, позвонок развивается как второй, осевой. Клетки как будто «думают», что они ближе к головной части зародыша. Кроме того, у мышей без генов Hoxa7 и Hoxb7, отвечающих за первые позвонки грудного отдела, позвонки, к которым должны крепиться ребра, развиваются как позвонки шеи, без ребер. Опять же, без этих генов клетки ведут себя так, как если бы они были ближе к голове, чем на самом деле. С другой стороны, когда удаляются Hoxa5 и Hoxa6, клетки в той же области тела «думают», что должны располагаться дальше от головы, где эти гены, естественно, дезактивируются. В результате седьмой позвонок шейного отдела имеет ребра, как полагается первому позвонку грудного отдела. То же происходит и ниже, если гены по правую сторону (см. рис. 28) удаляются.
В обеих системах сегментации, будь то «часы» и градиенты, благодаря которым тело делится на сомиты, или система HOX-генов, сообщающая клеткам этих сомитов специализацию, в масштабах организма протекают простые локальные процессы. В обоих случаях в этих процессах используются определенные временные промежутки для создания структур в пространстве. Это еще один пример того, как сложные клеточные системы эмбрионального развития пользуются простейшими математическими принципами и создают нечто гораздо большее, чем они сами.
Часть II
Постепенная детализация
Глава 7
Судьбоносные беседы
In principio erat verbum…[99]
Св. Иероним Стридонский
Процессы, описанные в главах 2–6, имеют внушительные последствия – бесформенное скопление клеток осталось далеко в прошлом. Теперь у эмбриона есть продольная ось тела с головным и хвостовым концами, дорсально расположенная нервная трубка и вентрально расположенный кишечник. По обе стороны нервной трубки залегают сомиты, а снаружи эмбрион покрыт эктодермой. Более того, благодаря избирательной экспрессии HOX-генов поведение клеток эмбриона специфично для уровня, на котором они находятся. Тем не менее на этой стадии развития эмбрион отличается от взрослого человека примерно так же, как эскиз отличается от законченного полотна: контуры видны, но их еще предстоит проработать.
На стадии формирования сомитов общее количество типов клеток в организме невелико. Есть клетки эктодермы, клетки нервной трубки, клетки кишечника, клетки сомитов и некоторые другие, но это лишь малая часть типов клеток, имеющихся у взрослого человека. Клетки первичных тканей эмбриона должны поделиться на группы, а группам предстоит специализироваться и дать начало разным структурам: костям, сухожилиям, мышцам и кровеносным сосудам. Более того, клетки должны сделать это по определенному алгоритму. Когда эмбрион впервые столкнулся с проблемой превращения одинаковых клеток в клетки разных типов (то есть когда у него обособилась внутренняя клеточная масса и трофэктодерма; глава 3), он использовал естественную асимметрию среды: клетки, часть поверхности которых была обращена в свободное пространство, дали начало трофэктодерме. Клетки нервной трубки и сомитов тоже полагаются в процессе дифференциации на сигналы, связанные с асимметрией окружающей среды. К этому моменту, однако, большая часть поступающей информации связана уже не с геометрическими характеристиками (такими, как наличие у клеток свободной поверхности), а с сигнальными молекулами, выделяемыми другими тканями. С помощью этих молекул соседние ткани ведут судьбоносные беседы и, таким образом, провоцируют друг у друга дифференцировку многих типов клеток, взаимное расположение которых исключительно точно задано в пространстве.
Хороший пример этих «клеточных бесед» – дифференцировка специализированных зон нервной трубки, которые в дальнейшем дают начало разным частям нервной системы.[100] Нервная трубка, образованию которой была посвящена глава 5, проходит по средней линии эмбриона между эктодермой и нотохордом. Таким образом, нервная трубка оказывается естественным образом асимметричной среде: ее дорсальная[101] поверхность ближе к эктодерме, а вентральная – к нотохорду. Эта асимметричная близость к нотохорду очень важна, потому что его клетки секретируют белок, получивший название Sonic Hedgehog («ежик Соник»).[102] Концентрация этого белка меняется по градиенту: она максимальна вблизи нотохорда и снижается по мере удаления от него.[103] Клетки нервной трубки чувствительны к белку Sonic Hedgehog, и те из них, которые расположены ближе к нотохорду, стимулируются очень сильно (рис. 30). В результате эти клетки начинают синтезировать новые белки и отличаться от других клеток нервной трубки. С этого момента они называются клетками вентральной пластинки.