Описанная выше система решает проблему деления мезенхимы на серию сегментов. Это не решает проблему создания на основе сомитов структур, которые не одинаковы на разных уровнях передне-задней оси, скажем позвонков в организме человека. Сомиты в грудной области, например, специализируются на формировании позвонков среднего размера, к которым крепятся ребра. Сегменты поясничного отдела формируют крупные позвонки. К ним ребра уже не крепятся! Сомиты в области крестца формируют мелкие позвонки, которые сливаются воедино, чтобы образовать заднюю часть костяного кольца, которая защищает область таза. В связи с этим клетки каждого сомита должны «знать», в какой области организма они находятся. После многолетних генетических исследований ученые начинают догадываться, каким образом клетки это «понимают». По нашим нынешним соображениям, эта система использует время, переводя пространственную информацию с молекулярного уровня на уровень эмбриона как целого. Безусловно, по мере дальнейших исследований наши представления о работе этой системы могут и измениться.
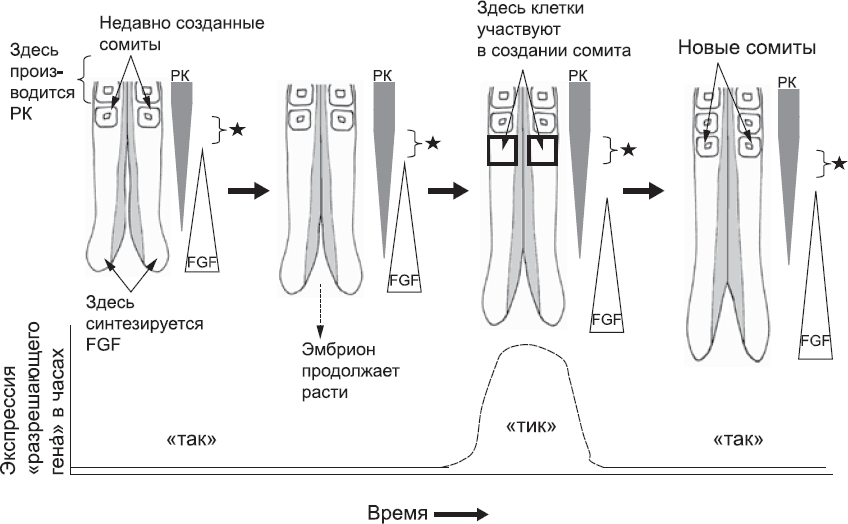
Рис. 27. Принятая в настоящее время модель использования времени и пространства для разделения непрерывного слоя ткани на сомиты. Диаграмма слева, как на рис. 24, отображает заднюю часть эмбриона сразу после формирования новой пары сомитов. Ретиноевая кислота (РК), синтезированная более ранними сомитами, распространяется к хвосту и постепенно расходуется. На диаграмме это показано заштрихованными перевернутыми треугольниками. Белок FGF, синтезируемый в хвостовой почке, распространяется к голове и также постепенно уходит (на диаграмме это отмечено треугольниками). Соотношение РК к FGF варьируется на разных участках оси «голова – хвост» мезенхимы вплоть до последних сомитов. Если клетки находятся в определенном диапазоне соотношения РК к FGF, они могут участвовать в формировании сомита. «Разрешающая зона» отмечена на схеме черной звездочкой. С течением времени рост хвостовой части эмбриона продолжается. Созревающие сомиты начинают синтезировать РК. Так «окно», определяющее клетки, которые будут создавать новый сомит, постепенно продвигается к «хвосту». Клетки в этой зоне возьмутся за формирование сомита только тогда, когда часы находятся в фазе «тик». К тому времени, когда случится следующий «тик», клетки, создававшие сомит, завершат свою работу. Так вырисовывается четкая граница между старыми и новыми сомитами. Затем процесс повторяется столько раз, сколько нужно, чтобы создать все сомиты тела
Необходимая пространственная информация хранится на молекулярном уровне в самой ДНК в виде четырех генных кластеров. Типов так называемых HOX-генов всего тринадцать. Их так и принято нумеровать: от 1 до 13. В каждом из четырех кластеров – HOXA, HOXB, HOXC и HOXD – HOX-гены, расположены в порядке нумерации (рис. 28). Таким образом, ген типа 1 в кластере HOXA называется HOXA1, ген типа 1 в кластере HOXB – HOXB1. Следовательно, в кластере HOXA гены располагаются в порядке HOXA1, A2, A3 и т. д. Ни в одном кластере нет версии каждого гена от 1 до 13. Таким образом, если положение генов изобразить на диаграмме, поместив гены одного типа в одну колонку, в каждом HOX-кластере будут пробелы (рис. 28). Есть весомые свидетельства того, что у наших далеких предков был только один HOX-кластер (именно этот кластер по-прежнему есть у насекомых, например у мучного хрущака), но в процессе эволюции позвоночных HOX-кластер был скопирован дважды. Так образовались первые два кластера (как у бесчелюстные позвоночных), а затем и еще два (как у челюстноротых позвоночных).[93] Это объясняет, почему HOXA1 так похож на HOXB1, а HOXA2 так напоминает HOXB2 и т. д. С тех пор каждый кластер утратил некоторые гены. Предположительно это случилось потому, что вскоре после дублирования гены были настолько похожи, что любой из них мог заменять другой. Потеря предками гипотетического H OX D 6, например, не имела значения, если H OX A 6, H OX B 6 и HOXC6 могли выполнять все его функции. Со временем однотипные гены разных кластеров мутировали и приобретали значительные функциональные отличия, а значит, дальнейшие потери стали невозможны. Верить или не верить таким историям об эволюции – решать вам. Факт в том, что человеческие HOX-кластеры теперь имеют такой вид (рис. 28).
Поразительно, что в течение каких-то 460 млн лет челюстноротые позвоночные плавали, ходили или летали, а порядок HOX-генов внутри кластера ни разу не менялся! При этом большинство других генов уже пережили огромное количество перемен и многократно меняли взаимное расположение. На сегодняшний день последовательность генов у разных типов животных отличается. Причины такого постоянства расположения HOX-генов в кластере напрямую связаны с порядком специализации сегментов вдоль оси тела.
Рис. 28. Генетическая структура четырех человеческих HOX-кластеров. Каждая горизонтальная линия обозначает непрерывную область хромосомы. Каждая ячейка с номером – ген HOX (например, ячейка 1 кластера HOXA – HOXA1 и т. д.)
Генные HOX-кластеры начинают проявлять активность в структурировании организма уже во время гаструляции. На каждом участке оси от шейного отдела к поясничному эта активность проявляется незадолго до образования сомитов. В то время как клетки в ходе гаструляции продвигаются через узелок, активируются гены с левого конца HOX-кластеров («левый» на рис. 28). Процесс начинается с будущего шейного к поясничному отделу. Позже, когда клетки, направленные на формирование задней части тела, вовлекаются в гаструляцию, активируются и гены, расположенные правее в HOX-кластерах. Еще через некоторое время, когда гаструляция продвинется дальше по оси «голова – хвост», активизируются гены, изображенные на рис. 28 правее. В каждом случае клетки «запоминают» набор HOX-генов, которые они «подключили», как только появились из узелка. Они в течение долгого промежутка времени поддерживают активность этих генов. Возможно, не стоило так упрощать, но в данном случае это даже полезно.
Таким образом, клетки, которые проходят стадию гаструляции раньше, «включают» HOX-гены слева (см. рис. 28). Клетки, подвергшиеся гаструляции позже, активируют HOX-гены справа. Как это можно контролировать? Есть способы. Можно направить процесс активации потенциальных генов направо через кластеры HOX-генов. Клетки в это время ожидают гаструляции. Согласно этой идее, когда клетка ныряет через узелок, чтобы гаструляция произошла, она на самом деле активирует экспрессию множества HOX-генов, которые потенциально могут быть задействованы в нужный момент. Она помнит этот набор. Клетки, которые гаструлируют позже, находятся дальше от будущей головы эмбриона, экспрессируют набор генов дальше вправо (см. рис. 28). Так волна активации потенциальных генов будет иметь больше времени для продвижения направо через кластер. По этому механизму, порядок генов в HOX-кластерах на молекулярном уровне транслируется на порядок экспрессии генов HOX на эмбриональном уровне. А эмбрион уже достиг миллиметрового диапазона. Это один из немногих примеров взаимосвязи генетической и эмбриональной структуры.
Прежде чем изучать последствия экспрессии генов на разных уровнях оси «голова – хвост», стоит потратить немного времени, чтобы выяснить, как волна активации потенциальных генов в первую очередь проходит через HOX-кластеры. Детали этого процесса до сих пор тщательно исследуются, но общая схема механизма, кажется, начинает вырисовываться сквозь дымку множества экспериментальных результатов. В хромосомах ДНК может быть довольно компактно упакована среди белков. Это полезно для экономии пространства, но при этом гены недоступны для транскрипции. Принципиально ДНК может быть и в свободной, легко доступной форме. Большую часть времени многие участки хромосом плотно упакованы, а более свободные «петли» ДНК выходят за их пределы. Некоторые участки упакованы плотно. Некоторые никогда не были так упакованы. Некоторые из них могут быть в любом состоянии, в зависимости от присутствия того или иного ДНК-связывающего белка. Последние данные, полученные в ходе экспериментов над мышиными эмбрионами, позволяют предположить, что по крайней мере один из hoх-кластеров,[94] hoxb, может быть в любом из обозначенных состояний.
