Лишенная резцов и окрашенная в неровный бурый цвет, левая половина нижней челюсти взрослого толсторога из Куртака имеет хорошую сохранность (рис. 3). На принадлежность челюсти барана к виду Ovis nivicola помимо небольших размеров в сравнении с таковой архара указывает наличие всего двух премоляров, наблюдающихся у 40–45 % рецептных толсторогов [Млекопитающие… 1963, с. 1063]. Размеры нижней челюсти снежного барана с левобережья Енисея следующие (мм): 1. Длина — 199. 2. Длина от заднего края до подбородочного отверстия — 162. 3. Альвеолярная длина зубного ряда — 79.4. Коронарная длина премоляров (Р3-Р4) -18,5. 5. Коронарная длина коренных зубов (М1-М3) — 47. 6. Высота нижней челюсти за Р 4— 30. 7. Высота нижней челюсти за М3 — 46. 8. Ширина основания восходящей ветви — 52. 9. Длина и ширина М 3— 29 и 11,5.
Foramen mandibulae и foramen mentale наеденной челюсти забиты сцементированной карбонатизированной массой, факт — косвенно свидетельствующий о существенной древности находки. Часть собранных здесь же среднеплейстоценовых (?) остатков крупных млекопитающих: волка, малого пещерного медведя, носорога, лошадей, мамонта, лося, благородного оленя и некоторых других видов заключены в известковистые конкреции. Этот признак по мнению Н. И. Дроздова, В. П. Чехи и Е. В. Артемьева [1992] позволяет предварительно сопоставить фаунистические находки с данного участка с «нижнеплейстоценовым аллювием погребенной долины» [там же, с. 88]. Но возможна и более молодая дата — средний, начало позднего плейстоцена.
Еще одна пясть снежного барана была обнаружена в местонахождении Усть-Обор, расположенном в среднем течении долины р. Хилок (Западное Забайкалье). Первые сборы ископаемых остатков млекопитающих проведены на Усть-Оборе в 1971 году археологом М. В. Константиновым, приславшим кости для определения в Институт археологии и этнографии (Новосибирск). По наблюдениям М. В. Константинова (личное сообщение): «Местонахождение представляет собой эрозионный уступ высотой 10–15 метров, рассеченный рядом оврагов. В одном из них я собрал сначала подъемный материал, а затем заметил костеносные линзы на глубине 5,25 м, из которых извлек ряд костей».
Из присланной небольшой коллекции Усть-Обора автору удалось кроме пястной кости и двух обломков плюсны снежного барана определить следующих млекопитающих: Panthera spelaea, Praeovibos sp., Equus ex gr. mosbachensis-germanicus, Coelodonta sp., Spirocerus sp..
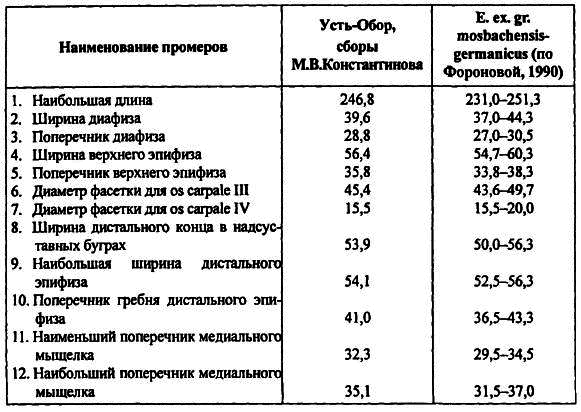
Поскольку до недавнего времени нижний возрастной предел снежного барана был определен как поздний плейстоцен [Каталог… 1981, с. 407], особый интерес представляет геологический возраст тафоценоза Усть-Обора, что вернее всего можно определить по найденному там же, к сожалению, единичному образцу пястной кости лошади.
Размеры и пропорции (по методике Айзенман) пястной кости лошади из Усть-Обора с очевидностью свидетельствуют о принадлежности этого животного к группе эквид, обозначаемой как Equus mosbachensis-germanicus и позволяют датировать пясть лошади и соответственно снежного барана средним плейстоценом, чему не противоречит и находка пястной костиPraeovibos sp. (табл. 3). Возможно, сходной по возрасту с усть-оборской находкой является какая-то часть скелета Ovis nivicola [ Боескоров2001] на реке Буотама в Центральной Якутии.
Морфологические особенности метаподий снежного барана
Теперь обратимся к некоторым морфологическим особенностям скелета конечностей снежного барана в связи с их ископаемыми находками. Известно, что для горно-скальных копытных в сравнении с равнинными животными характерна укороченность дистальных отделов ног [ Егоров1955]. К примеру, у современного снежного барана длина пясти по отношению к сумме длин плечевой и лучевой костей по моим промерам (n=6) составляет 38,9 — 40,1 — 41,4 %; соответственно плюсневая кость по отношению к сумме длин бедра и голени выражена цифрами 33,2 — 33,7 — 35,1 %. У архара (n=5), как обитателя более пологих склонов.
Таблица 3. Пясть лошади Equus sp.. из Усть-Обора.
Этот порядок соотношений выражается для передней конечности 44,4 — 46,0 -49,0 %; для задней конечности — 36,8 — 38,7 — 41,9 %. То есть, пропорции основных элементов, составляющих конечности, дают толсто рогу преимущество в силе, а архару в скорости передвижения, что для каждого вида оказывается выгодным при спасении от четвероногих хищников и, возможно, от древнего человека.
Обращаясь к таблице 2,видим, что ископаемые метакарпы толсторога в среднем на 10 % длиннее рецентных пястных костей этого вида (170,0 — 181,1 -189,5 мм против 152,3 — 163,0 — 171,1 мм) при относительно более широком диафизе (индекс 4:1), сохраняющем в сечении одинаковые с пястными костями современного снежного барана пропорции (индекс 5:4).
По соотношению длины метакарпа и размеров сечения его в срединной части различают среди мелких полорогих виды, приспособленные к обитанию в равнинных условиях, либо, напротив, — в биотопах с резко выраженным рельефом [ Егоров1955, табл. 3, 4 и 10]. Применяя этот подход к нашему материалу, получаем несколько парадоксальный результат. Отношение длины пясти к ширине диафиза дают разброс цифр для фоссильных метаподий толсторога окрестностей Красноярска и в Усть-Оборе: 8,2–8,5 — 8,6; для рецентных костей чубука из Корякского нагорья и низовьев Лены имеем соответственно 8,6–9,2 — 9,5 (у архара по Егорову: 9,45–10,3 — 10,9; у сибирского козерога: 6,65 — 7,45 — 8,16). То есть плейстоценовые толстороги, обитавшие на просторах Южной Сибири с относительно плавным рельефом, по некоторым пропорциям метакарпа были более приспособленными к передвижению по крутым скалистым склонам, чем его ближайший современный северо-восточный сородич. Аналогичный вывод можно сделать, обратившись к сравнимым пропорциям между длиной метакарпа и передне-задним диаметром диафиза этой кости (индекс 1:5, табл. 2). Тот же результат дает несколько более сложный индекс массивности пястных костей, а именно, отношение длины пясти к полусумме ширины и поперечника диафиза. Используя собственные данные и материалы, опубликованные О. В. Егоровым [1955, табл. 10], получим соответствующий последовательный ряд (%):

То есть, судя по пропорциям пястной кости, снежный плейстоценовый баран по своим качествам при передвижении по крутым склонам превосходил рецентные популяции чубука и приближался к современному сибирскому козерогу. Архар в этом отношении за десятки тысячелетий остался неизменен. На рецептном материале для указанных видов подобный вывод сделан по соотношению длины пястной кости и длины грудопоясничного отдела позвоночного столба [ Сутула1990, табл. 3].
Подвидовое деление современных азиатских снежных баранов в соответствии с их географической локализацией построено в большинстве своем на различиях в окрасе, формах, размерах роговых чехлов и роговых стержней (у самцов), а также некоторьгх деталей строения и размеров черепов. Единого мнения о количестве рецентных подвидов у специалистов нет. Существенные морфологические отличия некоторых элементов посткраниального скелета плейстоценовых толсторогов южной части Сибири от современных представителей этого вида убеждают в необходимости выделения первого на ископаемом посткраниальном материале для территории Сибири подвида снежного барана. Не исключено, что с накоплением нового более полноценного плейстоценового материала появится потребность выделения особого вида. Известная гипотеза заселения баранами Северной Америки через Берингийский мост с территории Азии в начале четвертичного периода пока остается гипотезой, поскольку детально не обоснована достаточным палеофаунистическим материалом.