Тот факт, что на врожденное поведение влияют генетически обусловленные изменения в структуре и функциях органов чувств, нервной системы и т. д., конечно, не доказывает, что наследование поведения можно объяснить воздействием одних лишь генетических факторов; он показывает только, что для нормального поведения необходимо нормальное тело. Подумайте снова об аналогии с радио: изменения в приемнике влияют на его работу, но это не доказывает, что музыка, которая исходит из громкоговорителя, рождается внутри самого приемника.
В сфере поведения биохимические, физиологические и анатомические изменения могут предотвратить появление зародышевых структур, и, следовательно, целые моторные поля могут быть не в состоянии действовать, или они могут оказывать разные количественные воздействия на движения, контролируемые этими полями. И действительно, исследования в области наследования фиксированных моделей действия показывают, что «нетрудно найти вариации, которые незначительно влияют на выполнение этого действия, но такая модель все же проявляется во вполне узнаваемой форме, если она проявляется вообще».[227]
Наследование моторных полей, возможно, зависит от факторов, которые уже обсуждались в связи с наследованием морфогенетических полей (глава 7). Вообще говоря, у гибридов между двумя породами или видами преобладание моторных полей одного над моторными полями другого, вероятно, зависит от относительной силы морфического резонанса со стороны родительских типов (ср. рис. 19). Если один принадлежит к хорошо установившимся породе или виду, а другой — к относительно новым, с малочисленной предшествующей популяцией, можно ожидать, что моторные поля первого будут доминировать. Но если родительские породы или виды утвердились одинаково хорошо, гибриды будут подпадать под влияние их обоих в одинаковой степени.
И это то, что происходит на самом деле. В некоторых случаях результаты принимают весьма причудливую форму, поскольку модели поведения родительских типов несовместимы друг с другом. Один из примеров дают гибриды, которые получаются при скрещивании двух видов неразлучников (небольших попугаев). Оба родительских вида строят свои гнезда из полосок, которые они вырывают из листьев одинаковым способом; но если один (неразлучник Фишера) несет потом эти полоски к гнезду в клюве, другой (неразлучник персиковоликий) переносит их, засовывая между своими перьями. Гибриды выдергивают полоски из листьев нормально, но затем ведут себя весьма странным образом, иногда засовывая полоски между перьев, иногда перенося их в клюве; но, даже когда они переносят их в клюве, они топорщат перья внизу спины и пытаются спрятать туда свои полоски.[228]
11.2. Морфический резонанс и поведение: экспериментальная проверка
В механистической биологии проводится резкое различие между врожденным и обученным поведением: предполагается, что первое «генетически запрограммировано», или «закодировано», в ДНК, тогда как последнее рассматривается как результат физико-химических изменений в нервной системе. Невозможно представить, каким способом эти изменения могут специфически модифицировать ДНК (такое требование выдвигали бы последователи ламаркизма); поэтому считается невозможным, чтобы обученное поведение, приобретенное животным, могло наследоваться его потомством (конечно, исключая «культурное наследование», при котором потомство воспринимает модели поведения от своих родителей).
В противоположность этому, согласно гипотезе формативной причинности, нет качественного различия между врожденным и обученным поведением, поскольку оба зависят от моторных полей, даваемых морфическим резонансом (раздел 10.1). Поэтому эта гипотеза допускает возможную передачу обученного поведения от одного животного к другому и приводит к проверяемым предсказаниям, которые отличаются не только от таковых ортодоксальной теории наследования, но также и от предсказаний ламаркизма.
Рассмотрим следующий эксперимент. Дикие животные помещаются в условия, где они учатся реагировать на данный стимул неким характерным образом. Затем их заставляют повторять эту модель поведения много раз. Согласно гипотезе, новое моторное поле будет усилено морфическим резонансом, который не только приведет к тому, что поведение обученных животных станет все более привычным, но и повлияет так же, хотя и менее специфично, на других животных, подвергаемых тому же стимулу: чем больше будет число животных, научившихся выполнять данное задание в прошлом, тем легче его будет выучить последующим подобным же животным. Поэтому в экспериментах такого типа должна быть возможность наблюдать прогрессивное увеличение скорости обучения не только у животных, происходящих от обученных предков, но также у генетически подобных животных, происходящих от необученных предков. Это предсказание отличается от такового ламаркистской теории, согласно которой только потомки обученных животных должны обучаться быстрее. А по общепринятой теории не должно наблюдаться увеличение скорости обучения потомков как необученных, так и обученных животных.
Сказанное можно суммировать следующим образом: повышенная скорость обучения в последовательных поколениях как тренированных, так и нетренированных линий подтверждала бы гипотезу формативной причинности: увеличение скорости только в тренированных линиях — ламаркизм, а отсутствие такого увеличения в обоих линиях — ортодоксальную теорию.
Эксперименты этого типа фактически уже проводились. Результаты подтверждают гипотезу формативной причинности.
Оригинальный эксперимент был начат в Гарварде в 1920 году У. Мак-Дугаллом, который надеялся провести тщательную проверку возможности наследования приобретенных особенностей поведения, предполагаемых ламаркизмом. Экспериментальными животными были белые крысы уистарской (Wistar) линии, которые — бережно выращивались в лабораторных условиях в течение многих поколений. Их задача состояла в том, чтобы научиться выбираться из специально сконструированного бака с водой, доплывая до одного из двух проходов, из которого можно было выйти наружу. Ложный проход был ярко освещен, тогда как истинный проход не освещался. Если крыса выходила через освещенный проход, она получала удар электрошоком. Два прохода освещались по очереди, один раз — первый, следующий раз — второй. За меру скорости обучения принималось число ошибок, которые допускала крыса, прежде чем она усваивала, что выходить надо через неосвещенный проход:
«Некоторым крысам требовалось целых 330 погружений, причем приблизительно половина из них сопровождалась электрошоком, прежде чем они научались избегать освещенный проход. Во всех случаях процесс обучения внезапно достигал критической точки. В течение длительного времени животное выказывало явное отвращение к освещенному проходу, часто колебалось, прежде чем войти в него, поворачивало назад или отчаянно бросалось внутрь; но, не уловив постоянной связи между ярким светом и шоком, оно продолжало устремляться в этот проход столь же часто, как и в другой. Наконец в обучении наступал момент, когда животные, оказавшись перед освещенным проходом, определенно и решительно поворачивали обратно, искали другой, смутно видимый проход и спокойно вылезали наружу. После достижения этой точки в обучении ни одно животное не делало ошибку, снова выбирая яркий проход, разве что в очень редких случаях».[229]
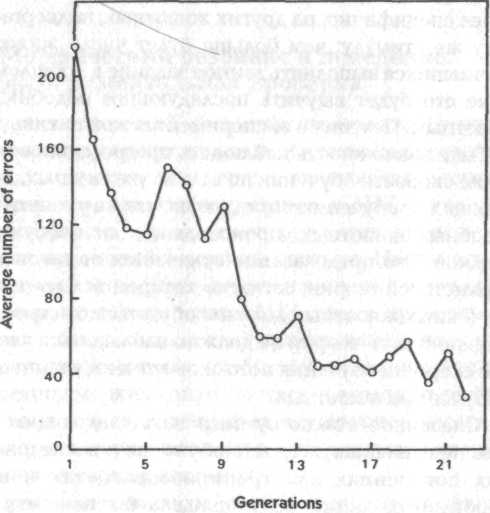
Рис. 28. Среднее число ошибок у последовательных поколений крыс, отобранных в каждом поколении по медленности обучения (данные из McDougall, 1938)
В каждом поколении крысы, от которых должно было быть выведено следующее поколение, отбирались случайным образом до того, как определялась скорость их обучения, тогда как спаривание происходило после их тестирования. Такая процедура была выбрана для того, чтобы избежать любой возможности сознательного или неосознанного отбора в пользу более быстро обучающихся крыс.