9.4. Повторяющийся морфогенез специализированных структур
Движения большинства животных зависят от изменения формы, скорее, неких специализированных структур, нежели тела в целом.
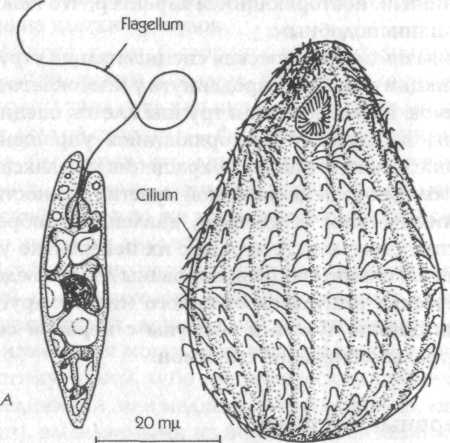
Рис. 25. А — жгутиковое, Euglena gracilis (Raven et al., 1976); В — простейшее, Tetmhymena pyriformis (Mackinnon and Hawes, 1961)
Многие одноклеточные организмы приводятся в движение за счет биения хлыстоподобных выростов, именуемых жгутиками, или ресничками, тогда как форма остальной части клетки остается более или менее неизменной (рис. 25). Эти подвижные органеллы содержат длинные трубчатые элементы, очень похожие на цитоплазматические микротрубочки; изменение формы белков, связанных с трубочками, порождает силу сдвига, под действием которой сгибаются жгутики, или реснички.[192]
У реснисчатых движения многих отдельных ресничек координируется таким образом, что волны биения проходят по поверхности клетки. У некоторых видов эта координация, по-видимому, зависит от механического воздействия ресничек на их соседей; а у других — от системы возбуждения внутри клетки, вероятно связанной с фибриллами, соединяющими основания ресничек.[193]
Если плывущее одноклеточное, например инфузория (Paramecium), встречает неблагоприятный стимул, направление биения ресничек изменяется на противоположное: организм отплывает назад, а затем снова плывет вперед в другом направлении.[194] Эта реакция избегания, возможно, запускается входом в клетку кальция или других ионов в результате изменения проницаемости мембраны, вызванного данным стимулом.[195]
Изменение формы бьющихся жгутиков, или ресничек, а также управление этим биением имеет такой стереотипный, повторяющийся характер, что кажется почти машиноподобным.
Эта квазимеханистическая специализация структуры и функции еще более продвинута у многоклеточных организмов, Целые клетки и группы клеток специализированы и проходят повторяющийся упрощенный морфогенез в своих циклах сокращения и релаксации; другие обладают специфической чувствительностью к свету, химическим веществам, давлению, вибрации или другим стимулам. А нервы, с их невероятно удлиненными аксонами, специализированы для проведения электрических импульсов от одного места к другому, соединяя органы чувств и мускулы с нервной сетью или центральной нервной системой.
9.5. Нервные системы
Так же как биение отдельной реснички на поверхности одноклеточного животного координировано с таковым соседних одноклеточных с помощью определенных физических связей, сокращение индивидуальных клеток мускулов координировано с помощью определяющих импульсов, проходящих по нервам. Когда несколько соседних клеток активируются одним нервом, их можно побудить сократиться одновременно. А если активность нерва является частью системы управления высшего уровня, сокращение различных групп клеток может координироваться ритмично, как это происходит с мускулом, который сохраняет напряжение в течение определенного периода времени. Далее, системы еще более высоких уровней контролируют повторяющиеся циклы сокращения различных мускулов, например в ногах животного, когда оно бежит. Таким образом, иерархически организованная деятельность нервной системы позволяет осуществлять координацию по ступеням, что было бы невозможно, если бы поля, управляющие движениями организмов, действовали непосредственно на клетки мускулов.
Но хотя, с одной стороны, нервы функционируют детерминистским образом, передавая определенные импульсы типа «все или ничего» из одного места в другое, с другой стороны — формативная причинность не могла бы управлять движениями животных через нервную систему, если бы деятельности нервов не был свойствен в то же время вероятностный характер. И это на самом деле так.
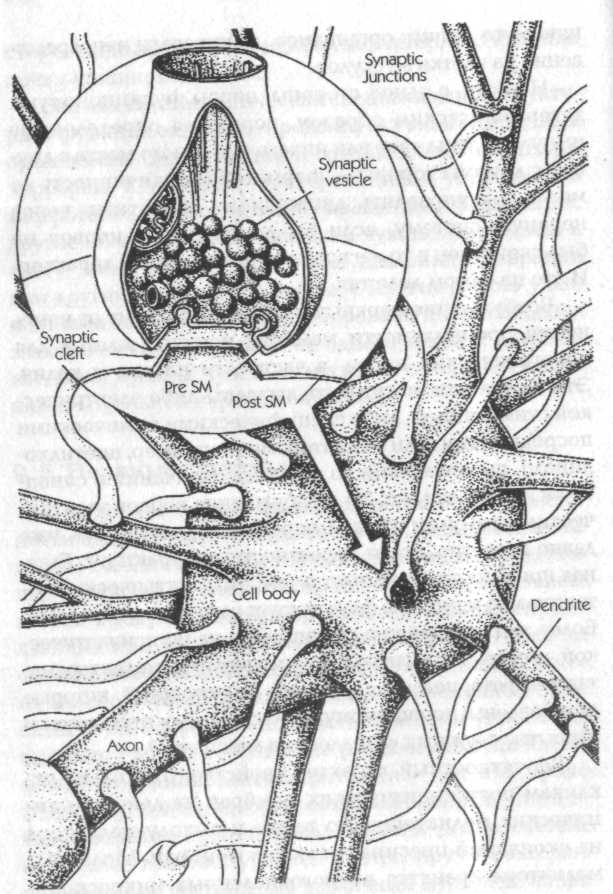
Рис. 26. Часть нервной клетки с множеством синапсов на поверхности. На вставке крупным планом показан один синапс. Pre SM — пресинаптическая мембрана;
Post SM — постсинаптическая мембрана (Kristic, 1979)
Возбуждение нервных импульсов зависит от изменений проницаемости мембран нервных клеток для неорганических ионов, в частности натрия и калия. Эти изменения могут быть вызваны либо электрической стимуляцией, либо специфическими химическими посредниками, или медиаторами (например, ацетилхолином), выделяемыми из нервных окончаний в синаптических узлах {рис. 26). Возбуждение нервов электрическим стимулом вблизи порогового уровня, как уже давно известно, носит вероятностный характер.[196] Главная причина этого состоит в том, что электрический потенциал на мембране флуктуирует случайным образом.[197] Более того, изменения потенциалов на постсинаптической мембране, вызванные химическими медиаторами, также претерпевают случайные флуктуации,[198] которые обусловлены, по-видимому, вероятностным открытием и закрытием ионных «каналов» на мембране.[199]
Вероятностный характер свойствен не только откликам постсинаптических мембран на действие химических медиаторов, но также и выходу медиатора из окончаний пресинаптических мембран. Молекулы медиатора хранятся в многочисленных микроскопических пузырьках (рис. 26) и выделяются в синаптическую щель при слиянии этих пузырьков с мембраной. Этот процесс происходит самопроизвольно через случайные интервалы времени, вызывая так называемые миниатюрные потенциалы концевой пластинки. Скорость секреции сильно возрастает, когда на нервное окончание поступает импульс, но и тогда опять-таки слияние везикул с мембраной происходит вероятностным образом.[200]
В мозгу типичная нервная клетка имеет тысячи тонких нитеобразных выростов, которые оканчиваются синаптическими контактами с другими нервными клетками, и, обратно, отростки сотен или тысяч других нервных клеток оканчиваются синапсами на поверхности данной клетки (рис. 26). Некоторые из этих нервных окончаний выделяют возбуждающие вещества (трансмиттеры), которые способствуют «возгоранию» импульса; другие оказывают ингибирующее действие и уменьшают тенденцию нерва к возбуждению. Включение импульсов фактически зависит от баланса между возбуждающими и ингибирующими воздействиями со стороны сотен синапсов. Кажется вероятным, что в любой данный момент времени во многих нервных клетках мозга этот баланс находится на столь критическом уровне, что возбуждение происходит или не происходит в результате вероятностных флуктуации в клеточных мембранах синапсов.
Таким образом, детерминистское распространение нервных импульсов с одного участка тела на другой связано с высокой степенью неопределенности (индетерминизма) в центральной нервной системе, которая, по настоящей гипотезе, упорядочена и организована под действием формативной причинности.