Глава 10. Инстинкт и обучение
10.1. Влияние прошлых действий
Подобно морфогенетическим полям, моторные поля задаются морфическим резонансом от прошлых подобных систем. Детали строения животного и организация колебательной активности его нервной системы обычно подобны таковым у него самого более, нежели у какого-либо другого животного. Таким образом, наиболее специфическим морфическим резонансом, действующим на данное животное, будет резонанс от его собственной формы в прошлом (ср. раздел 6.5). Следующий наиболее специфический резонанс будет от генетически подобных животных, которые жили в тех же условиях, и наименее специфический — от животных других пород, живущих в других условиях. В «долинной» модели хреоды {рис. 5] последний стабилизирует общую форму, тогда как более специфический резонанс определяет детали топологии дна этой долины.
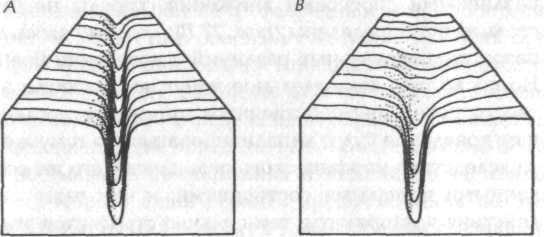
Рис. 27. Схематическое изображение глубоко канализированной хреоды (А) и хреоды, слабо канализированной на начальных стадиях (В)
«Контуры» долины хреод зависят от степени подобия между поведением родственных животных той же породы или вида. Если их модели движения варьируются слабо, морфический резонанс порождает глубокие и узкие хреоды, представленные долинами с крутыми краями {рис. 27 А). Такие хреоды оказывают сильное канализирующее действие на поведение последующих индивидуумов, которые по этой причине будут стремиться вести себя весьма сходным образом. Стереотипные модели поведения, обусловленные такими хреодами, на низших уровнях проявляются как рефлексы, а на высших уровнях — как инстинкты.
С другой стороны, если подобные друг другу животные достигают конечных форм своих моторных полей различными способами движения, хреоды не будут столь четко определены {рис. 27 В); поэтому здесь диапазон индивидуальных различий в поведении больше. Но после того как отдельное животное достигло цели своего поведения собственным путем, его последующее поведение будет канализироваться по тому же пути вследствие морфического резонанса с его же собственными прошлыми состояниями; и чем чаще такие действия повторяются, тем сильнее становится эта канализация. Такие характерные индивидуальные хреоды проявляют себя как привычки.
Таким образом, с точки зрения гипотезы формативной причинности между инстинктами и привычками разница лишь в степени: и те и другие зависят от морфического резонанса, первые — с бесчисленными предшествовавшими индивидуумами того же вида, а вторые — преимущественно с прошлыми состояниями того же индивидуума.
Этим мы не хотим сказать, что рефлексы и инстинкты не зависят от весьма специфически организованного морфогенеза нервной системы. Очевидно, что такая зависимость существует. Мы не хотим также сказать, что в процессах обучения не происходят физические или химические изменения в нервной системе, которые облегчают повторение данного вида движения. Возможно, в простой нервной системе, осуществляющей стереотипные функции, потенциал для таких изменений может быть уже «встроен» в систему «проводов» таким образом, что обучение происходит квазимеханически. Например, было обнаружено, что у улитки Aplysia строение нервной системы почти одинаково у разных особей, вплоть до мельчайших деталей расположения возбуждающих и ингибирующих синапсов на отдельных клетках. Очень простые виды обучения встречаются в связи с рефлекторным втягиванием жабер во впадину под покровной пластинкой, а именно привыкание к безвредным и приобретение чувствительности к опасным стимулам; при этих процессах функционирование отдельных возбуждающих и ингибирующих синапсов, действующих на индивидуальные клетки, определенным образом изменяется.[206] Конечно, простое описание этих процессов само по себе не выявляет причин таких изменений; в настоящее время можно лишь строить предположения на этот счет. Одно из них состоит в том, что эти модификации химического происхождения и связаны, вероятно, с изменениями в фосфорилировании белков.[207] Но как возникла эта тонкая специализация структуры и функций нервов и синапсов? Проблема переносится в область морфогенеза.
Нервные системы высших животных значительно больше варьируются от индивидуума к индивидууму, чем у беспозвоночных типа Aplysia, и они гораздо сложнее. Очень мало известно о том, каким образом сохраняются заученные способы поведения,[208] но накоплено уже достаточно сведений, чтобы утверждать, что здесь не может быть простого объяснения на уровне специфически локализованных физических и химических «следов» в нервной ткани.
Многочисленные исследования показали, что у млекопитающих привычки, образовавшиеся при обучении, часто сохраняются после значительного повреждения коры и участков подкорки головного мозга. Более того, когда происходит потеря памяти, она не является тесно связанной с местонахождением таких повреждений, но зависит, скорее, от общего количества поврежденной ткани. К. С. Лэшли суммировал результаты сотен экспериментов следующим образом:
«Невозможно продемонстрировать изолированный участок как след памяти где-либо в нервной системе. Ограниченные области могут быть существенными для обучения или сохранения определенной активности, но в таких областях их части в функциональном отношении эквивалентны».[209]
Подобный же феномен был продемонстрирован на беспозвоночном — осьминоге: наблюдения сохранения приобретенных привычек после разрушения различных частей вертикальной доли мозга привели к кажущемуся парадоксальным выводу, что «память находится везде одновременно и нигде в частности».[210]
С механистической точки зрения эти результаты сильно озадачивают. В попытках найти им объяснение было высказано предположение, что «следы» памяти каким-то образом распределены в мозгу способом, аналогичным тому, который используется при сохранении информации в виде интерференционных картин в голограмме.[211] Но это пока не более чем неопределенное соображение.
Гипотеза формативной причинности дает альтернативное объяснение, в котором сохранение приобретенных привычек, несмотря на повреждение мозга, выглядит гораздо менее загадочно: привычки зависят от моторных полей, которые вообще не сохраняются в мозгу, но приходят непосредственно от его прошлого состояния путем морфического резонанса.
Некоторые приложения гипотезы формативной причинности применительно к проблемам инстинкта и обучения рассматриваются в следующих разделах, а в главе 11 предлагаются способы, с помощью которых можно экспериментально отличить предсказания, вытекающие из этой гипотезы, от предсказаний механистической теории.