Ответы на эти стимулы включают сложные физико-химические изменения в клетках и тканях, а в некоторых случаях зависят от различий в распределении гормонов, таких как ауксин. Однако эти реакции не могут быть объяснены с помощью одних только физико-химических изменений, но могут быть поняты лишь с привлечением общих морфогенетических полей растений. Например, благодаря присущей им полярности растения производят на одном конце стебли, на другом — корни. Направленный стимул гравитации руководит этим поляризованным развитием так, что стебли растут вверх, а корни вниз. Действие гравитационного поля на зерна крахмала в клетках и соответствующие изменения в распределении гормонов действительно являются причиной этих ориентированных движений роста, но не могут сами по себе объяснить существование полярности; не объясняют они и тот факт, что главные стебли и корни реагируют диаметрально противоположным образом, а также различные особенности роста деревьев, трав, вьющихся и ползучих растений и особенности разветвления стебля и корневой системы различных видов. Все эти признаки зависят от морфогенетических полей.
Несмотря на то что большая часть движений растений происходит только в молодых растущих органах, некоторые структуры сохраняют способность двигаться даже тогда, когда они вполне созрели, например, цветы, которые открываются и закрываются каждый день, и листья, складывающиеся ночью. На эти движения влияют интенсивность света и другие факторы среды; они находятся также под контролем «физиологических часов» и продолжают происходить приблизительно раз в сутки, даже если растения помещаются в неизменяемые условия.[187] Листья или лепестки открываются, потому что специализированные клетки в «шарнирной» (hinge) области у их основания разбухают; и они закрываются, когда эти клетки теряют воду вследствие изменения проницаемости их мембран по отношению к неорганическим ионам.[188] Восстановление тургорного давления является активным процессом, требующим затрат энергии и сравнимым с ростом.
Помимо «сонных» движений листья некоторых растений движутся в течение дня в ответ на изменение положения солнца. Например, у голубиного гороха, Cajanus cajan, листочки, на которые падает солнце, ориентированы приблизительно параллельно солнечным лучам, что минимизирует площадь поверхности, подверженной интенсивному тропическому излучению. Но листья в тени ориентируются под прямыми углами к падающим лучам, тем самым улавливая максимальное количество света. Эти реакции зависят от направления и интенсивности света, падающего на специализированные узлы в листьях (листовые подушечки). В течение дня листья и листочки непрерывно приспосабливают свое положение к движению солнца по небу. Ночью они принимают свое вертикальное «спящее» положение: листовые подушечки чувствительны к гравитации так же, как и к свету.
Рис. 23. Листья чувствительного растения, Mimosa pudica. Слева — не стимулированные; справа — стимулированные
У «чувствительного» растения, Mimosa pudica, листочки закрываются и листья смотрят вниз ночью, как и у многих других бобовых растений. Но эти движения быстро происходят также и в дневное время в ответ на механический стимул {рис. 23). Стимул вызывает распространение вниз по листу волны электрической деполяризации, подобной нервному импульсу; если стимул достаточно сильный, импульс распространяется на другие листья, которые также свертываются.[189] Подобно этому, у растения венерина мухоловка, Dionaea muscipula, механическая стимуляция чувствительных волосков на поверхности листа вызывает движение электрического импульса к разбухшим «шарнирным» клеткам, которые быстро теряют воду; в результате лист захлопывается как ловушка вокруг несчастных насекомых, которые затем перевариваются.[190]
Эти движения листьев и листочков в ответ на действие света, силы тяжести и механическую стимуляцию возможны потому, что специализированные клетки способны терять воду, а затем вырастать снова; следовательно, они сохранили упрощенный морфогенетический потенциал, тогда как у большинства других тканей он теряется, когда они созревают и перестают расти. Обратимые движения этих специализированных структур можно рассматривать как предельные случаи морфогенеза, в которых изменения формы стали стереотипными и повторяющимися. Но их квазимеханистическая простота в эволюционном отношении является вторичной, а не первичной: она возникла из ранее существовавшей предпосылки, в которой чувствительность к стимулам среды связана с ростом и морфогенезом растения в целом.
9.3. Амебоидное движение
Амебы движутся с помощью объемного тока своей цитоплазмы в вырастающие выступы — псевдоподии. В норме они перемещаются по поверхности твердых объектов посредством непрерывного растягивания своих передних концов. Но если до псевдоподий дотронуться или если они соприкасаются с теплом или концентрированными растворами химических веществ, они перестают расти, вместо них вырастают другие, и тогда клетка меняет направление движения. Если новые псевдоподии снова встречают какие-либо потенциально опасные стимулы, они также останавливаются, и амеба уходит от них в другом направлении. Эта система «проб и ошибок» действует до тех пор, пока они не находят путь без препятствий или неблагоприятных стимулов.[191]
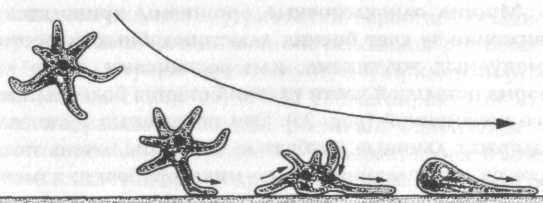
Рис. 24. Метод, с помощью которого плавающая амеба переходит на твердую поверхность (согласно данным, приведенным в Jennings, 1906)
У свободно плавающих амеб, не испытывающих действия какого-либо особого направленного стимула, нет соответствующей ориентации роста; псевдоподии продолжают развиваться в разных направлениях, пока одна из них не соприкоснется с поверхностью, вдоль которой она может ползти {рис. 24).
Вытягивание псевдоподий происходит предположительно под влиянием специфического поляризованного морфогенетического поля. Ориентация, в которой начинают формироваться новые псевдоподии, может в большой степени зависеть от случайных флуктуации внутри клетки; виртуальные псевдоподии, выдвинутые из тела клетки, затем актуализируются посредством организации сократительных волокон и других структур в цитоплазме. Этот процесс продолжается до тех пор, пока развитие псевдоподий не подавляется стимулами из окружения или в результате конкуренции с псевдоподиями, растущими в других направлениях.
Возможно, тот факт, что амебоидные движения зависят от непрерывных морфогенетических процессов, засвидетельствован в самом названии Amoeba proteus в виде намека на мифическое морское божество, которое непрерывно изменяло свой вид.
Амебы питаются, поглощая частицы еды, такие как бактерии, путем фагоцитоза: псевдоподии растут вокруг частицы, соприкасающейся с поверхностью клетки; мембраны псевдоподий сливаются, и частица оказывается заключенной внутри клетки и окруженной частью клеточной мембраны. Другие мембранные пузырьки (везикулы), содержащие пищеварительные ферменты, сливаются с фагоцитарной везикулой, и еда переваривается. Этот тип морфогенеза отличается от характерного для передвижения клетки и осуществляется предположительно под действием другого морфогенетического поля, ориентация которого зависит от контакта потенциальной питательной частицы с мембраной клетки. Частица, соприкасающаяся с мембраной, может рассматриваться как морфогенетический зародыш; конечной формой здесь является частица, поглощенная клеткой. Хреода фагоцитоза, приводящая к этой конечной форме, задается морфическим резонансом от всех подобных актов фагоцитоза, происходивших в прошлом у тех же амеб.