Ч. с. получили широкое применение в экспериментах на ускорителях заряженных частиц , т.к. они позволяют выделять частицы, скорость которых заключена в определённом интервале. С ростом энергии ускорителей и, следовательно, с ростом энергии частиц особенно широкое применение получили газовые Ч. с., обладающие способностью выделять частицы ультрарелятивистских энергий, для которых (1— b) << 1. Угол излучения J в газе очень мал, мала и интенсивность излучения на единицу пути. Чтобы получить вспышку света, достаточную для регистрации, приходится увеличивать длину газовых Ч. с. до 10 м и более. В газовых Ч. с. можно плавно менять показатель преломления, изменяя давление рабочего газа.
Ч. с. существуют 3 типов: пороговые, дифференциальные и счётчики полного поглощения. Основными характеристиками первых 2 типов Ч. с. являются эффективность регистрации и разрешающая способность по скорости частиц, т. е. способность счётчика разделять две частицы, двигающиеся с близкими скоростями. Пороговый Ч. с. должен регистрировать все частицы со скоростями, большими некоторой (пороговой), поэтому оптическая система такого Ч. с. (комбинация линз и зеркал) должна собрать, по возможности, весь излученный свет на катод ФЭУ.
Дифференциальные Ч. с. регистрируют частицы, движущиеся в некотором интервале скоростей от u1 до u2 . В традиционных дифференциальных Ч. с. это достигается выделением оптической системой света, излучаемого в интервале соответствующих углов от J1 до J2 . Линза или сферическое зеркало, помещенное на пути черенкового света, фокусирует свет, излученный под углом J, в кольцо с радиусом
R = f J, (4)
где f ¾ фокусное расстояние линзы или зеркала. Если в фокусе системы поместить щелевую кольцевую диафрагму, а за диафрагмой один или несколько ФЭУ, то в такой системе свет будет зарегистрирован только для частиц, излучающих свет в определённом интервале углов. В дифференциальных Ч. с. с прецизионной оптической системой можно выделить частицы, скорость которых отличается всего на 10¾6 от скорости др. частиц. Такие Ч. с. требуют особого контроля давления газа и формирования параллельного пучка частиц.
Ч. с. полного поглощения предназначены для регистрации и спектрометрии электронов и g-квантов. В отличие от рассмотренных Ч. с., где частица теряла в радиаторе ничтожно малую долю энергии, Ч. с. полного поглощения содержит блок радиаторов большой толщины, в котором электрон или g-квант образует электронно-фотонную лавину и теряет всю или большую часть своей энергии. Как правило, радиаторы в этом случае изготавливают из стекла с большим содержанием свинца. В радиаторе из такого стекла, например толщиной 40 см , может практически полностью тормозиться электрон с энергией до 10 Гэв. Количество света, излучаемого в Ч. с. полного поглощения, пропорционально энергии первичного электрона или g-кванта. Разрешающая способность DE Ч. с. полного поглощения (по энергии) зависит от энергии и для самых чувствительных ФЭУ может быть выражена формулой:
%
где E — энергия электрона в Гэв.
Лит.: Джелли Дж., Черенковское излучение и его применения, пер. с англ., М., 1960; Зрелов В. П., Излучение Вавилова—Черенкова и его применение в физике высоких энергий, ч. 1¾2, М., 1968.
В. С. Кафтанов.
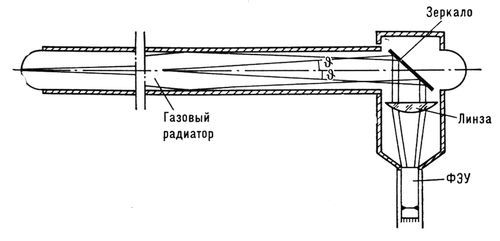
Схема газового порогового черенковского счётчика на 70 Гэв ускорителя Института физики высоких энергий (СССР). Черенковский свет собирается на катод ФЭУ с помощью оптической системы, состоящей из плоского зеркала и кварцевой линзы.
Черенок
Черено'к, часть растения, используемая для вегетативного размножения . Ч. заготавливают с высококачественных растений (называемых маточными, или материнскими). Образовавшиеся из Ч. растения сохраняют свойства и признаки маточных. Различают Ч. корневые, стеблевые и листовые. При определённых условиях выращивания на стеблевых Ч. образуются корни, на корневых — почки, на листовых — и почки и корни. Способность растений к размножению Ч. зависит от видовых и сортовых особенностей маточных растений, а также от условий внешней среды, в которых происходит укоренение Ч. (температура, влажность, аэрация и т.д.). См. также ст. Черенкование .
Череп
Че'реп (cranium), скелет головы позвоночных животных и человека. Различают осевой и висцеральный Ч. Осевой, или мозговой, Ч. представляет переднее продолжение осевого скелета туловища, разрастающееся вокруг головного мозга, органов обоняния и внутреннего уха. Висцеральный, или лицевой, Ч. — скелет переднего отдела кишечника (глотки), первоначально представленный жаберными (висцеральными) дугами, разделяющими жаберные щели.
Череп животных. Изменения Ч. в процессе эволюционного развития организмов обусловлены прогрессивным развитием головного мозга и органов чувств, заменой жаберного дыхания лёгочным и сменой различных способов питания, что было связано с переходом организмов из водной среды на сушу. Осевой Ч. состоит из мозговой коробки (вмещающей головной мозг), носовых капсул (окружающих органы обоняния) и ушных капсул (заключающих внутреннее ухо). Мозговая коробка подразделяется на передний (прехордальный) и задний (хордальный) отделы. Прехордальный отдел объединяет глазничную и носовую области Ч., хордальный — затылочную и ушную. Прехордальный отдел развивается из трабекулярных хрящей и расположенных над ними глазничных (орбитальных) хрящей (рис. 1 ). У эмбрионов хордальный отдел развивается вокруг головного конца хорды из расположенных по его сторонам парахордальных хрящей и ушных капсул. Парахордалии соответствуют слившимся невральным дугам наиболее передних позвонков, что позволяет говорить о «позвоночном» происхождении хордального отдела. У кистепёрых рыб оба отдела остаются самостоятельными, у остальных позвоночных они срастаются у эмбрионов. Прехордальный отдел вмещает обычно только передний мозг (большие полушария), хордальный — большую часть головного мозга. На Ч. взрослых животных границу между отделами обозначают по линии гипофиза и отверстия для выхода из Ч. тройничного нерва. Носовые капсулы срастаются с прехордальным отделом. У кистепёрых рыб и наземных позвоночных помимо наружных ноздрей имеются внутренние (хоаны), открывающиеся в переднюю часть ротовой полости.
В зависимости от строения глазничного отдела различают платибазальный и тропибазальный типы строения мозговой коробки (рис. 2 ). В платибазальном черепе прехордальный отдел имеет широкое основание, головной мозг проникает в глазничную область (круглоротые, акуловые и двоякодышащие рыбы, современные земноводные, млекопитающие). В тропибазальном черепе прехордальный отдел имеет узкое основание, большие полушария расположены позади глазничной области (остальные позвоночные).
Висцеральный Ч. служит опорой для передней части пищеварительного тракта. Он состоит из метамерно расположенных хрящевых дуг, охватывающих подобно обручам начальный отдел пищеварительного тракта. У всех позвоночных, исключая наиболее примитивных — бесчелюстных, передние висцеральные дуги преобразованы в челюсти . Хрящевая основа верхней челюсти — нёбноквадратный хрящ сформировался в результате слияния верхних отделов двух передних дуг — предчелюстной и собственно челюстной. Хрящевая основа нижней челюсти — меккелев хрящ — соответствует нижнему отделу второй висцеральной (челюстной) дуги. Верхний отдел третьей висцеральной (гиоидной) дуги у рыб превращен в т. н. подвесок, обычно участвующий в прикреплении челюстей к осевому Ч. У большинства рыб челюстная дуга сочленяется с осевым Ч. только посредством подвеска (гиостилия). У химер и двоякодышащих рыб, а также у наземных позвоночных нёбноквадратный хрящ непосредственно соединяется с осевым Ч. без участия подвеска (аутостилия). У вымерших панцирных и примитивных костных рыб и у некоторых акул челюстная дуга сочленяется с осевым Ч. как непосредственно, так и с помощью подвеска (амфистилия) (рис. 3 ). У наземных позвоночных подвесок превращается в слуховую косточку, проводящую звуки от барабанной перепонки к внутреннему уху, задние висцеральные (собственно жаберные) дуги преобразуются в подъязычную кость и хрящи гортани. У млекопитающих появляются дополнительные слуховые косточки — наковальня и молоточек, сформировавшиеся из отчленившихся задних элементов нёбноквадратного и меккелева хрящей — квадратной и сочленовной костей.