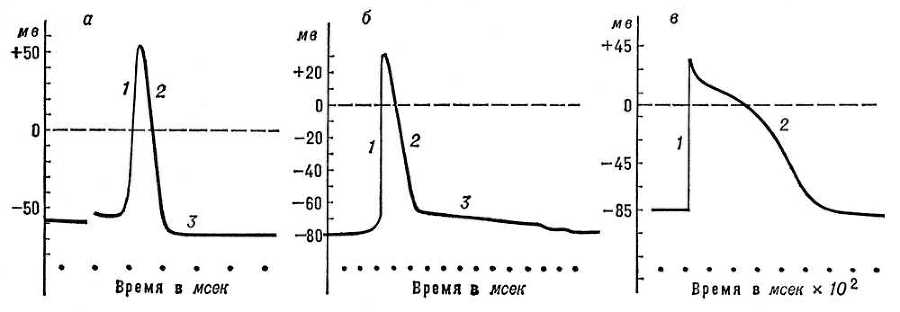
Амплитуда ПД большинства нервных и мышечных волокон примерно одинакова: 110—120 мв. Длительность ПД варьирует в широких пределах: у теплокровных животных длительность ПД нервных волокон, наиболее быстро проводящих возбуждение, — 0,3—0,4 мсек, у волокон же мышц сердца — 50—600 мсек. В растительных клетках пресноводной водоросли хара ПД продолжается около 20 сек. Характерной особенностью ПД, отличающей его от других форм ответа клетки на раздражение, является то, что он подчиняется правилу «всё или ничего», т. е. возникает только при достижении раздражителем некоторого порогового значения, и дальнейшее увеличение интенсивности раздражителя уже не сказывается ни на амплитуде, ни на продолжительности ПД. Потенциал действия — один из важнейших компонентов процесса возбуждения. В нервных волокнах он обеспечивает проведение возбуждения от чувствительных окончаний (рецепторов) к телу нервной клетки и от неё — к синаптическим окончаниям (см. Синапсы), расположенным на различных нервных, мышечных или железистых клетках. Поступая в эффекторные окончания, ПД вызывает выделение (секрецию) определённой порции специфических химических веществ, т. н. медиаторов, оказывающих возбуждающее или тормозящее влияние на соответствующие клетки. В мышечных волокнах распространяющийся ПД вызывает цепь физико-химических реакций, лежащих в основе процесса сокращения мышц. Проведение ПД вдоль нервных и мышечных волокон осуществляется т. н. локальными токами, или токами действия, возникающими между возбуждённым (деполяризованным) и соседними с ним покоящимися участками мембраны (см. Возбуждение). Токи действия регистрируются обычными внеклеточными электродами; при этом кривая имеет двухфазный характер: первая фаза соответствует приходу ПД под ближний электрод, вторая — под дальний электрод (рис. 4).
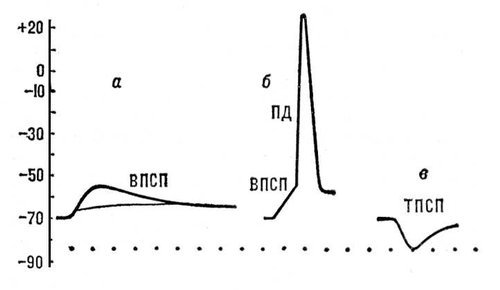
Постсинаптические потенциалы (ПСП) возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мв и длительность 10—15 мсек. ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД (рис. 5, а, б). ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора (рис. 5, в). В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки.
Генераторные потенциалы возникают в мембране чувствительных нервных окончаний — рецепторов. Они внешне сходны с ВПСП — их амплитуда порядка нескольких мв и зависит от силы приложенного к рецептору раздражения (рис. 6). Когда генераторный потенциал достигает порогового (критического) значения, в соседнем участке мембраны нервного волокна возникает распространяющийся ПД. Ионный механизм генераторных потенциалов ещё недостаточно изучен.
Наряду с перечисленными относительно быстро развивающимися Б. п., в нервных клетках, волокнах гладких мышц и некоторых растительных клетках регистрируются также очень медленные колебания мембранного потенциала неизвестной природы, причём на гребне волны деполяризации мембраны часто возникают разряды импульсов.
Все Б. п. могут быть зарегистрированы и точно измерены только с помощью внутриклеточных микроэлектродов, позволяющих отводить разности потенциалов между внутренней и наружной сторонами мембраны клетки. При отведении колебаний Б. п. от целых нервов, мышц или мозга с помощью поверхностных электродов регистрируется лишь суммарно потенциал множества синхронно или, чаще, асинхронно работающих клеток. Так, электромиограмма представляет собой результат сложения (интерференции) ПД множества скелетных мышечных волокон; электрокардиограмма — результирующая колебаний электрических потенциалов мышечных волокон различных отделов сердца; электроэнцефалограмма — результат суммации главным образом ВПСП и ТПСП множества клеток различных слоев коры больших полушарий. Регистрация таких интерференционных электрограмм, хотя и не позволяет анализировать колебания Б. п. отдельных клеток, имеет важное значение для суждения о состоянии исследуемого органа в целом. В клинической практике электромиограмму, электрокардиограмму и электроэнцефалограмму регистрируют с помощью электродов, расположенных на коже соответствующих частей тела. Оценка данных, полученных этими методами, основана на сопоставлении изменений характера соответствующей кривой с результатами клинических, физиологических и патологоанатомических исследований.
Лит.; Физиология человека, М., 1966; Гальвани Л. и Вольта А., Избр. работы о животном электричестве, М. — Л., 1937; Ходжкин А., Нервный импульс, пер. с англ., М., 1965; Экклс Дж., Физиология нервных клеток, пер. с англ., М., 1959; его же, Физиология синапсов, М., 1966; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969.
Б. И. Ходоров.
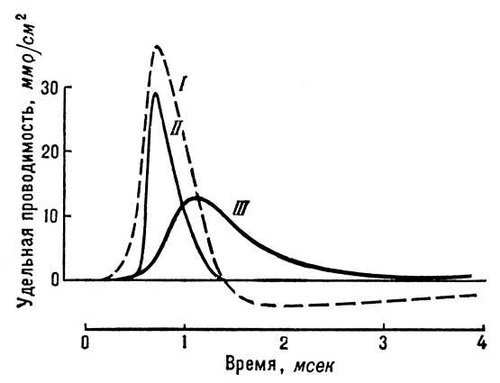
Рис. 3. Изменения натриевой и калиевой проводимости мембраны нервного волокна во время генерации потенциала действия (I). Изменения проводимости пропорциональны изменениям проницаемости для Na+(II) и К+(III).
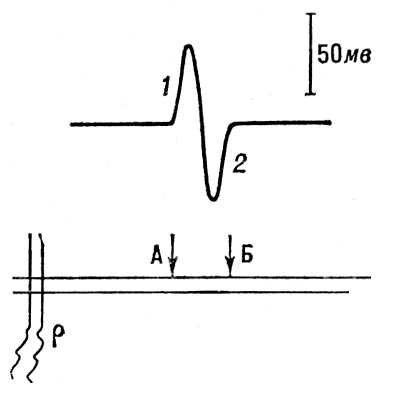
Рис. 4. Регистрация распространения потенциала действия вдоль нервного волокна. А, Б — внеклеточные электроды; р — раздражающие электроды. Вверху — отклонение луча осциллографа под влиянием волны возбуждения; 1 — волна возбуждения под электродом А; 2 — под электродом Б.
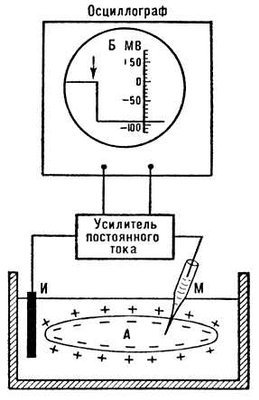
Рис. 1. Схема измерений мембранного потенциала покоя с помощью внутриклеточного стеклянного микроэлектрода (М). Второй электрод (И) помещен в омывающую клетку жидкость.
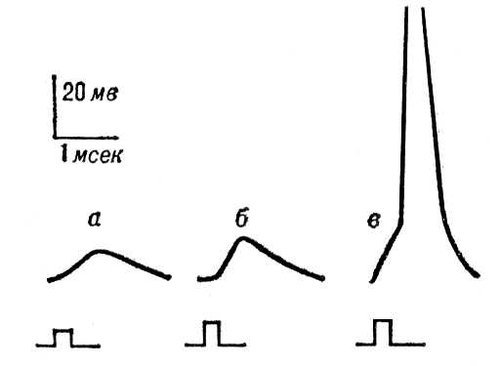
Рис. 6. Генераторные потенциалы: увеличение амплитуды при усилении раздражения рецептора (а — в). При достижении порогового значения (в) генераторный потенциал вызвал в чувствительном нервном волокне потенциал действия.
Рис. 5. Постсинаптические потенциалы: а — подпороговые ВПСП, возникающие в нервной клетке в ответ на раздражения соответствующих нервных волокон; б — ВПСП, достигший порогового значения, достаточного для возникновения ПД; в — ТПСП, вызванный раздражением тормозных нервных волокон.
Рис. 2. Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов: а — гигантского аксона кальмара; б — скелетного мышечного волокна; в — волокна мышцы сердца собаки; 1 — восходящая фаза ПД; 2 — нисходящая фаза; 3 — следовая гиперполяризация (а) и следовая деполяризация (б).
Биоэлектрические явления
Биоэлектри'ческиеявле'ния, см. Биоэлектрические потенциалы.
Биоэнергетика
Биоэнерге'тика, биологическая энергетика, изучает механизмы преобразования энергии в процессах жизнедеятельности организмов. Иначе говоря, Б. рассматривает явления жизнедеятельности в их энергетическом аспекте. Методы и подходы к изучаемым явлениям, применяемые в Б., — физико-химические, объекты и задачи — биологические. Т. о., Б. стоит на стыке этих наук и является частью молекулярной биологии,биофизики и биохимии.