В 2017 году Всемирный экономический форум представил новый Индекс инклюзивного развития, основанный на ряде ключевых показателей эффективности (KPI) и позволяющий провести всестороннюю оценку стандартов жизни не только на текущем уровне развития, но и с учетом показателей за прошедшие пять лет (World Economic Forum, 2017). Индекс развития человеческого потенциала и Индекс инклюзивного развития во многом пересекаются: их рейтинг совпадает в шести из 10 ведущих стран (Норвегия, Швейцария, Исландия, Дания, Нидерланды, Австралия). Пожалуй, наиболее интересными новыми расчетами является количественное выражение счастья или удовлетворенности жизнью.
О маленьком государстве Бутан, расположенном в Гималаях, заговорили в 1972 году, когда его четвертый король Джигме Сингье Вангчук предложил измерять развитие королевства с помощью индекса под названием Валовое национальное счастье (GNH Centre, 2016). Идея, конечно, привлекательная, но превратить яркий образ в математически надежный и регулярно отслеживаемый индикатор – совсем другое дело. Во всяком случае, для США периода после Второй мировой войны у нас имеются достаточно убедительные доказательства того, что счастье не является функциональной переменной роста. Институт Гэллапа с 1948 года проводит нерегулярные опросы американцев о том, насколько счастливыми они себя ощущают (Carroll, 2007). В 1948 году очень счастливыми ощущали себя 43 % американцев. Пик показателя, 55 %, был достигнут в 2004 году, а самое низкое значение пришлось на опрос после событий 11 сентября 2001 года – 37 %. Но к 2006 году оно поднялось до 49 %, то есть едва изменилось по сравнению с цифрами полувековой давности (47 % в 1952 году)!
Удовлетворенность жизнью тесно связана с рядом качественных преимуществ, которые нелегко отразить с помощью простых, самых распространенных, количественных единиц. Питание и жилье являются двумя лучшими примерами этой реальности. Как бы важно ни было отслеживать рост средней ежедневной доступности пищевой энергии на душу населения, цифры могут демонстрировать обманчиво обнадеживающую картину. С улучшением питания к нему стали предъявляться повышенные требования, выходящие за рамки удовлетворения энергетических потребностей: пища может обеспечивать достаточное количество углеводов и жиров и удовлетворять минимальному уровню высококачественного белка, но быть бедной микроэлементами (витаминами и минералами). Следует отметить, что низкий уровень потребления фруктов и овощей (основных источников микроэлементов) считается основным фактором, повышающим риск хронических заболеваний, но Сигел и др. (Siegel et al., 2014) продемонстрировали, что в большинстве стран их потребление ниже рекомендуемого. В 2009 году глобальная нехватка составила 22 % с медианным соотношением между обеспечением и потребностью на уровне 0,42 в странах с низким доходом и 1,02 в обеспеченных странах.
В начале Нового времени развитие научных методов исследований и изобретение и применение новых, мощных математических и аналитических инструментов (математического анализа в середине XVII века, успехов в области теоретической физики и химии и основ современной экономики и демографии в течение XIX века) позволили анализировать рост в чисто количественной форме и использовать релевантные формулы роста для прогноза долгосрочных траекторий изучаемых феноменов. Роберт Мальтус (1766–1834), один из основоположников демографических и экономических исследований, вызвал серьезную обеспокоенность своими выводами, в которых противопоставлялся линейный рост средств к существованию и экспоненциальный рост населения (Malthus, 1798).
В отличие от Мальтуса, Пьер Франсуа Ферхюльст (1804–1849), бельгийский математик, сегодня известен только историкам науки, статистикам, демографам и биологам. Но через четыре десятка лет после публикации работы Мальтуса Ферхюльст внес существенный вклад в наше понимание роста, опубликовав первые реалистичные формулы, разработанные специально для того, чтобы выразить развитие ограниченного роста (Verhulst, 1838; 1845; 1847). Подобный рост управляет не только развитием всех организмов, но и повышением производительности новых методов организации труда, распространением множества инноваций и внедрением множества потребительских продуктов. Прежде чем начать раскрывать тему феноменов роста и их траекторий (в главе 2), я предложу краткое, но вполне исчерпывающее введение в характер этих формальных моделей роста и соответствующих им кривых.
Линейный и экспоненциальный рост
Это две распространенные, но совершенно различные формы роста, траектории которых отражают простые равенства. «Относительно медленный и устойчивый» будет лучшим качественным описанием линейного роста, а «ускоряющийся и переходящий в стремительный» – экспоненциального. Все, что подчиняется линейному росту, возрастает на одну ту же величину в течение заданного периода времени, следовательно, формула линейного роста выглядит просто:
Nt = N0 + kt,
где новое значение величины Nt (в момент времени t) рассчитывается путем увеличения начального значения (N0) на постоянную величину k за период времени t.
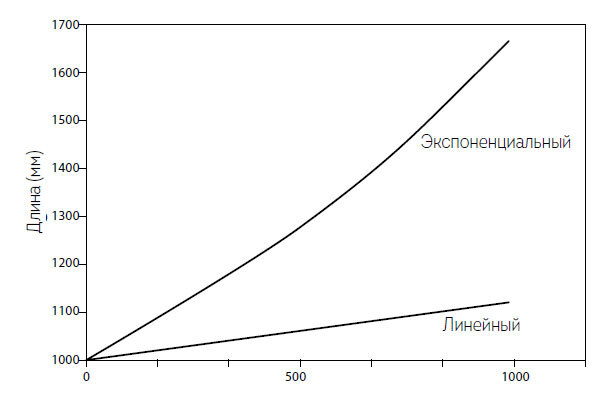
Анализ большого числа сталагмитов показывает, что эти конусообразные колонны солей кальция, образующиеся на полу пещер благодаря капающей воде, часто растут тысячелетиями почти линейно (White and Culver, 2012). Даже сравнительно быстрый рост со скоростью 0,1 мм в год означает, что сталагмит высотой 1 м вырастет за тысячу лет всего на 10 см (1000 мм + 1000 × 0,1). Если нанести этот результат на график, мы увидим плавно восходящую линию (рис. 1.3). Это, конечно, означает, что темп роста как доля общей высоты сталагмита будет постоянно снижаться. Для сталагмита, растущего со скоростью 0,1 мм в год в течение 1000 лет, он будет составлять 0,01 % в течение первого года, но всего 0,009 % спустя тысячелетие.
Рис. 1.3. Тысячелетие прироста сталагмитов, иллюстрирующее траектории линейного и экспоненциального роста
Для сравнения во всех случаях экспоненциального роста значение увеличивается в одинаковое число раз за каждый одинаковый период времени. Основной функциональной зависимостью является
Nt = N0 (1 + r)t,
где r – скорость роста, выраженная как доля единицы роста на единицу времени, например, при росте 7 % на единицу времени r = 0,07.
Экспоненциальный рост также можно выразить – после простой поправки на выбор единиц измерения времени – как
Nt = N0ert,
где e (e = 2,7183, основа натурального логарифма) возводится в степень rt, что легко проделать с помощью любого научного калькулятора. Мы можем представить себе пещеру, где количество капающей воды, содержащей одинаковую долю растворенных солей, постоянно возрастает, ведя к экспоненциальному росту сталагмита.
Если предположить очень малый прирост длины в размере 0,05 % в год, то сталагмит за 1000 лет увеличился бы в длину почти на 65 см (1,000 мм × 2,7180,0005 × 1000 = 1648,6 мм общей длины, или прирост в размере 64,86 см), что почти на 50 % больше, чем при линейном росте. Экспоненциальный рост отображается в виде восходящей кривой, крутизна подъема которой определяется скоростью роста (рис. 1.3). Через 10 000 лет линейно растущий сталагмит удвоил бы свою высоту, и она достигла бы 2 м, в то время как экспоненциально растущему сталагмиту понадобилась бы гигантская пещера, так как его высота составила бы 148,3 м. Экспонента – произведение скорости роста и времени, поэтому прирост может быть одинаково большим как в случае низкого прироста на более длинных интервалах времени, так и в случае более коротких интервалов более быстрого роста.
Еще одно простое сравнение показывает, что траектории линейного и экспоненциального роста находятся близко друг к другу на самых ранних стадиях роста, когда значения скорости роста и временного интервала невелики по сравнению с единицей: вскоре они начинают расходиться и в конце концов оказываются далеко друг от друга. Голд (Gold, 1992) считал, что колонии бактерий, живущих глубоко под землей, заполняют до 1 % всего пористого пространства в верхних 5 км земной коры, в то время как, по мнению Уитмана и др. (Whitman et al., 1998), объем, занимаемый микроорганизмами, составляет всего 0,016 % пористого пространства. Это все равно означает огромную совокупную массу микроорганизмов, но с крайне низким темпом размножения. Давайте предположим (ради простого примера), что физические и химические ограничения позволяют крошечной колонии, состоящей из 100 клеток (внезапно оказавшейся в результате сейсмического явления в новой полости скальной породы), расти на пять клеток в час. Очевидно, что к концу первого часа клеток будет 105, через 10 часов такого линейного роста колония достигнет 150 клеток, а через 50 и 100 часов – 350 и 600 соответственно.