Примеры расшифровки посредством феногеографии исторических событий важны как показатели огромных возможностей феногеографического анализа вообще.
Можно также привести примеры, не связанные с деятельностью человека, — естественных эволюционных процессов и их феногеографической расшифровки.
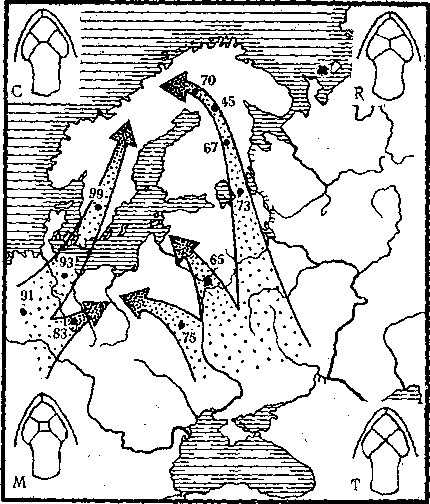
Финские исследователи изучали распространение четырех фенов сочетания предлобных щитков у живородящей ящерицы (рис. 15). По частоте проявлений разных фенов выяснилось следующее: финские популяции сходны с карельскими и восточноевропейскими, а шведские — с центрально- и западноевропейскими. Такая феногеографическая картина легко расшифровывается в свете сравнительно недавних исторических событий: около 11 тыс. лет назад вся Фенноскандия была под сплошным покровом ледника. По мере таяния ледника и потепления происходило заселение этой территории ящерицами. Фены показывают, что заселение шло из двух разных источников: первого — из Западной и Центральной Европы через существовавший тогда сухопутный перешеек на месте балтийских проливов, а второго — из каких-то центров на юго-востоке Европы, где сохранился этот вид.
Следующий пример значительно меньшего масштаба и касается сопоставления по четырем биохимическим фенам нескольких популяций мышевидных хомячков. Изученные популяции обитают на островах Мексиканского залива и на прилежащих частях материка. На островах фенофонд оказался очень однородным — здесь встречается только один фен из четырех, характерных для этой части ареала. На основании этих данных можно сделать два предположения. Первое предположение говорит о том, что все четыре популяции близки генетически между собой, они все происходят от одного «корня» — небольшой группы, когда-то попавшей на острова.
Откуда могли попасть эти исходные формы на острова? По сходству генофонда хомячков можно сделать вывод, что заселение островов шло не со стороны ближайших прибрежных популяций, а с довольно далеко отстоящей ныне от берега группы популяций, для которых характерно преобладание того же фена. Это — один из возможных ответов. Есть и другой. Набор фенов, или фенооблик, островных популяций и ближайших популяций, от которых они произошли, может отличаться потому, что среди немногих особей (основателей островной популяции) случайно не оказалось представителей, несущих остальные три признака.
Рис. 15. Различные сочетания (С, R, М, T) предлобных щитков у живородящей ящерицы и концентрация формы «М» в европейских популяциях этого вида.
Стрелками показаны предполагаемые пути заселения, цифрами — концентрация формы «М» в %
Второе предположение основано на том, что своеобразный фенооблик островных популяций может быть обусловлен естественным отбором, направленным против носителей иных биохимических аллелей, кроме одного-единственного, обладающего повышенной жизнеспособностью в данных условиях.
Все эти предположения доступны дальнейшей проверке. Надо дополнительно сравнить популяции по другим фенам. Совпадение по одному-двум фенам может быть случайным, но вероятность совпадения по нескольким фенам ничтожно мала и ею можно пренебречь. Другой путь проверки — сравнение условий существования островных популяций как с отдаленной от берега популяцией, сходной по биохимической характеристике, так и с какими-либо другими популяциями на этих и других островах, где сходное направление естественного отбора. Так можно из уравнения с несколькими неизвестными постепенно прийти к желанному уравнению с одним-двумя неизвестными.
Интересный пример касается гавайских дрозофил. (Научная статья американского генетика X. Карсона, посвященная их описанию, называется «Хромосомные следы происхождения видов».) Исследовались частота и характер расположения поперечных дисков (темных участков) в пяти хромосомах 69 видов гавайских дрозофил. Эти типичные дискретные признаки, касающиеся тонкой морфологии хромосом, можно считать морфологическими фенами. Их преимущество в том, что на пути от гена к признаку они находятся «близко» к генам. Все исследованные виды по исчерченности хромосом распределились на три группы, каждая из них включала филогенетически близкие формы. Эти группы, судя по фенетике, происходят от предковых форм, обитавших на о-ве Мауи. Поскольку геологическая история отдельных островов изучена достаточно хорошо, можно в данном случае как бы восстановить ход эволюционного процесса на протяжении нескольких сот тысяч лет. Примененный автором способ сравнения позволил высказать предположение, что несколько видов дрозофил, ныне обитающих на Гавайях, происходят от единственной самки: об этом говорит повторяющееся у всех уникальное сочетание дисков, которое не могло возникнуть независимо у разных особей.
Исследования гавайских дрозофил выполнены, несомненно, под влиянием классических работ по североамериканским дрозофилам, проведенных Ф. Г. Добржанским и А. Стертевантом в 30-х годах. В них показано, что, зная порядок генов в одной из хромосом, можно восстановить порядок и последовательность возникновения серии последовательных инверсий (поворотов на 180° участков внутри хромосомы), каждая из которых характерна для отдельных популяций и групп популяций. Эта работа остается лучшей в мировой литературе по точности и однозначности результатов. Она была выполнена не на фенетическом, а на генетическом уровне — все этапы ее подтверждались многочисленными экспериментами по скрещиванию дрозофил разных популяций.
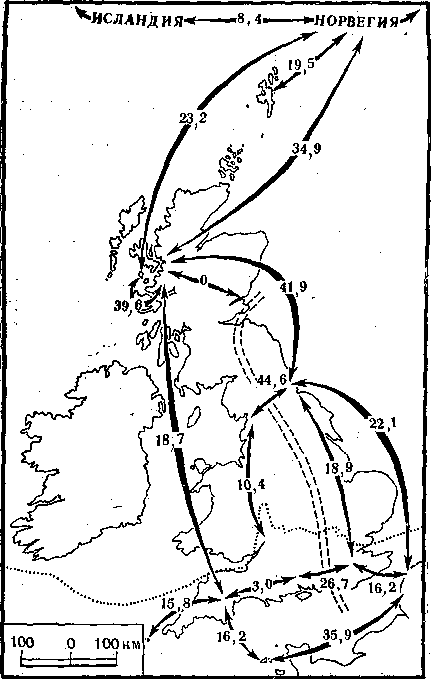
Рис. 16. Фенетические дистанции между популяциями лесных мышей разных районов Великобритании и соседних стран (в условных единицах)
Пунктиром обозначена граница максимального оледенения, двойным пунктиром — современный ареал доледниковых популяций
Фенетические данные (не столь надежные порознь, как генетические) в совокупности позволяют прийти к хорошо обоснованным микроэволюционным выводам.
Последним из серии примеров, иллюстрирующих феногеографический подход для расшифровки микрофилогенеза, будет пример исследования популяций лесных мышей в Англии. Это одно из наиболее обстоятельных феногеографических исследований было выполнено генетиком Р. Берри. Он изучал 20 фенов черепа у лесных мышей, обитающих в Шотландии и на прилежащих островах. По суммарному коэффициенту сходства была восстановлена история заселения этих островов. История состояла из двух периодов. Первый начался с момента максимального оледенения Европы, когда только самая южная часть современной Англии оставалась свободной ото льда (рис. 16). Именно здесь пережили ледниковое время немногочисленные тогда популяции мышей. По мере отступления ледника на север они стали распространяться к северу, но заняли только юго-восточную часть Англии.
Позднее, через несколько тысяч лет после этих событий, начался второй период истории. На север Шотландии уже в VIII в. приплыли викинги из Норвегии (их путь можно проследить по частоте фенов в популяции кошек), с ними сюда попали и норвежские лесные мыши. Постоянным центром активности викингов стал небольшой островок Ейгг среди Внутренних Гебридских островов. Так показывают и археологические исследования, и фенетические сравнения. Отсюда впоследствии и распространились лесные мыши во всей северной части Англии. Сейчас на большей части страны живут лесные мыши, происходящие из Норвегии; лишь на юго-востоке обитают потомки коренных доледниковых популяций.
Реконструкция микрофилогенезов большого числа видов позволит получить много интересных данных по особенностям протекания процесса микроэволюции и откроет новые возможности для глубокого понимания особенностей тех или иных видов. Конечно, такая реконструкция будет особенно убедительна при учете достаточно большой совокупности фенов.
Феногеография, структура вида и систематика
Феногеография, несомненно, поможет и в решении некоторых проблем внутривидовой таксономии.