Подробно рассмотрим только один пример плейотропии, связанный с геном карликовости у мышей. Это рецессивный ген, определяющий развитие животных, в два-три раза меньше нормальных размеров; он возник в одном из питомников белых мышей в 1929 г. Еще до остановки роста мышей-карликов можно отличить по целому ряду признаков: тупым мордочкам, коротким ушам и хвостикам, вялости, робости, чувствительности к колебаниям температуры и другим признакам. Продолжительность жизни карликовых мышей короче: и самцы и самки стерильны.
Этот яркий пример плейотропии в действии гена интересен одной особенностью. Оказалось, что карликовых мышей можно превратить в нормальных, если хирургически вживлять в их тело (под кожу) кусочки гипофиза крысы. (Гипофиз — небольшая железа внутренней секреции, расположенная в основании мозга и регулирующая выделение практически всех гормонов.) После ряда таких, операций карликовые животные преображались: они достигали нормальных размеров и начинали вести себя как нормальные мыши (правда, самки оставались стерильными).
Этот пример показывает, как наследственно определенные признаки могут резко измениться под влиянием среды, внешних условий. Значит, если выразиться точнее, не аллель карликовости ведет к стерильности, ранней остановке роста, стойкости к голоду и повышенной чувствительности к холоду, а действие аллеля карликовости ведет к такому изменению нормы реакции, при котором в обычных условиях развивается стерильность, рано прекращается рост, повышается стойкость к голоду и чувствительность к изменениям температуры.
Рассмотренный пример показывает также, что действие внешних по отношению к генотипу агентов способно замаскировать то или иное проявление гена. Эту возможную маскировку фенов, так же как и противоположный случай — возникновение псевдофенов — дискретных признаков в результате каких-то внешних вмешательств в развитие (например, травматические изменения), нужно постоянно иметь в виду. Такая всегда существующая неопределенность в выделении фенов — та цена, которую исследователю приходится платить природе при отказе от изучения каждого признака генетически, с применением методов скрещиваний в чреде поколений. Однако преувеличивать значение этой неопределенности для фенетического анализа не следует. Далее будет рассказано о приемах, позволяющих ослабить ее влияние при анализе конкретных данных.
Пока нет сколько-нибудь надежного сравнительного материала, позволяющего количественно оценить маскировку разных генов проявлением одного и того же фена. Однако, поскольку число признаков фенотипа практически бесконечно, а число генов у каждого вида имеет конечный характер, в будущем, при развитии фенетических методов анализа фенотипа, всегда можно найти фены, маркирующие ген либо непосредственно и однозначно, либо посредством сочетания. Но это — дело будущего.
Важнейшее значение для понимания природы фенов имеют работы английского генетика Г. Грюнеберга, проведенные в 50—60-х годах. Ученый сравнивал разные линии мышей (максимально однородные генетически в результате близкородственного разведения) по встречаемости мельчайших вариантов в строении скелета. Этими вариантами было развитие какого-либо отростка на костях, прохождение группы кровеносных сосудов в данном месте скелета через одно большое или несколько малых отверстий, расположение определенных отверстий для прохождения кровеносных сосудов и нервных стволов, различные аномалии зубной системы и т. п.
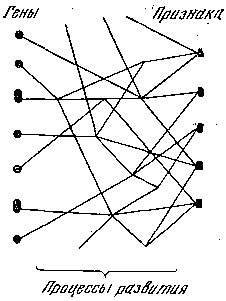
Рис. 5. Уточненная схема взаимоотношений гена и фена
Оказалось, что все варианты относились к числу пороговых: как только в процессе эмбрионального развития тот или иной зачаток будущей структуры достигал определенной величины, осуществлялось формирование будущей структуры — возникал отросток на кости, большое отверстие разделялось перегородкой надвое и т. п. Если же зачаток структуры в эмбриогенезе не достигал этой величины, подобные признаки не появлялись у взрослого организма. Хотя на величину зачатка оказывали влияние многочисленные взаимодействующие гены и внешние факторы, решающей причиной в конце концов приходилось считать генетическую. Изученные линии мышей хорошо различались по частотам таких фенов, и эти различия передавались по наследству.
Теперь, после более подробного рассмотрения связи гена и фена становится ясно, что изображенная на рис. 4 схема соотношения между генами и признаками нуждается в уточнении. Эти соотношения оказываются более сложными: плейотропия и полимерия представляют собой не свойство самих генов, а являются итогом различных реакций (от простейших биохимических до сложнейших морфогенетических), управляемых генами (рис. 5). Фены представляют собой видимые, улавливаемые наблюдателем звенья таких реакций.
Обычно в качестве фенов описываются конечные звенья морфогенетических цепей — «конечные продукты действия генов», по выражению австрийских исследователей Бергеля и Наса (1976). Однако дискретными признаками характеризуются и отдельные этапы в процессе индивидуального развития организма. Такой подход приложим к фенам у растений, как показано недавно работами по фенетике растений дагестанского исследователя М. Магомедмирзаева (1976–1977), который в изменчивости травянистых растений и деревьев выделяет элементарные, дискретные процессы, так называемые кванты морфогенеза.
После всего сказанного выше можно дать более полную формулировку понятия «фен». Феном называются любые дискретные альтернативные вариации признаков и свойств живых организмов, которые на всем имеющемся материале (обязательно многочисленном) далее неподразделимы без потери качества. Фены всегда отражают определенные черты генетической конституции данной особи, а своей частотой — генетическую структуру популяции и других (как более, так и менее крупных) групп особей данного вида.
Как найти фены?
В генетике для анализа того или иного признака достаточно скрестить пару особей и по характеру распределения признаков в потомстве (на протяжении ряда поколений) точно определить особенности любого признака или свойства. Для фенетического анализа необходимо значительное число особей. Без них исследователь не сможет составить достаточно полное представление о существующих в природе дискретных вариациях, среди которых и приходится искать фены.
Опыт, накопленный при исследовании фенов в разных группах животных и растений, позволяет предложить следующие пути выделения фенов на материале природных популяций.
1. Сопоставление наблюдаемой изменчивости по отдельным комплексам признаков или свойствам (окраске, рисунку, форме отдельных частей тела, вариантам строения отдельных органов и систем и т. п.).
2. Выделение признаков, подверженных возрастным и половым изменениям. Для первичного анализа они сложны, хотя среди них тоже можно найти фены, но это — более специальная задача. Обычно такие признаки не включаются в фенетическое исследование на животных, хотя, как упоминалось выше, могут быть с большим успехом использованы у растений.
3. Выделение в общей изменчивости признаков и свойств дискретных характеристик, их анализ на дальнейшую делимость (дробимость, подразделяемость).
Предположим, что изучаются такие особенности окраски, как рисунок и цвет. Дискретность в изменчивости нужно искать именно в цветовой гамме, а не в микроскопически определяемых характеристиках пигмента или микроструктуре поверхности органа, вызывающего цветовые эффекты (как у чешуекрылых). Равным образом было бы неправильно свести рисунок окраски (полосы, пятна на отдельных участках тела и т. п.) к изучению, например, гистологии кожного покрова. И в том и в другом случае исследователь потеряет качество изучаемой группы признаков и перейдет к изучению совсем другой группы признаков и свойств (среди которых тоже можно искать качественные альтернативные дискретные признаки).