Многие рыбы, обитающие в придонных слоях (галозавры, долгохвосты, ателеопы, ошибневые), независимо приобрели очень сходный тип строения, для которого характерны выступающее рыло, нижний рот, очень длинная, постепенно сходящая на нет хвостовая часть туловища, несущая по нижнему краю многолучевой аномальный плавник. Ундуляции такого плавника переводят рыбу в наклонное (головой вниз) положение, очень удобное для поиска корма на грунте. Все эти рыбы обладают плавучестью, близкой к нейтральной, как, впрочем, и ведущие сходный образ жизни придонные колючие акулы, но конвергентно сходный результат достигается принципиально разными путями — все перечисленные костистые рыбы имеют плавательный пузырь, а хрящевые регулируют плавучесть высоким содержанием легкого жира в огромной печени.
Скудность палеонтологического материала, значительные в ряде случаев отличия океанических таксонов рыб от родственных прибрежных форм, а также их сходство между собой, определяемое конвергенциями и параллелизмами, создают определенные трудности для воссоздания эволюционной истории рыб открытого океана. Тем не менее происхождение рецептной океанической ихтиофауны вполне очевидно: исходным материалом для ее формирования были шельфовые и неритические предковые формы. Заселение придонных глубин и открытой пелагиали, осуществлявшееся многими группами независимо друг от друга, представляется очень растянутым во времени. Датировка начала этого процесса, теряющаяся в далеком прошлом, невозможна без малооправданных спекуляций, а на примере эпипелагиали и мезобентали видно, что заселение основных океанских биотопов продолжается и поныне. В результате для каждого из этих биотопов характерна разновозрастная фауна.
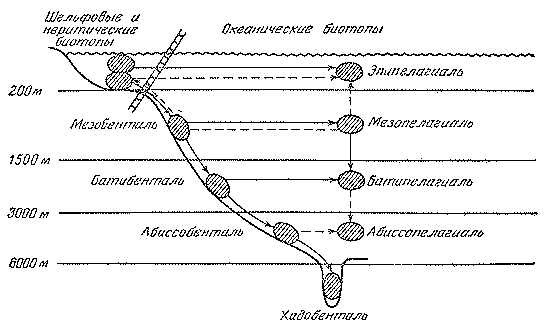
Рис. 44. Пути заселения океанических биотопов рыбами
Сплошными и штриховыми стрелками обозначены главные и второстепенные направления колонизации
Как известно, степень морфо-экологической специализации к условиям глубин, косвенно отражающая филогенетический возраст таксонов, была положена А. П. Андрияшевым в основу разделения глубоководных рыб на две группировки — древнеглубоководную и вторичноглубоководную. Такое разделение, несомненно реальное и очень наглядное в некоторых крайних случаях, все же довольно относительно и не дает возможности альтернативно определить принадлежность любой и каждой систематической группы рыб к одной из этих категорий. Представляется, что наиболее резко обособлены от ближайших родственников таксоны, возникшие в результате пелагизации, т. е. при переходе от бентического образа жизни к бентопелагическому, а затем к обитанию в безбрежной толще вод. Эти переходы носившие прогрессивный характер, сопровождались повышением общего уровня организации и соответствовали, таким образом, введенному академиком А. Н. Северцовым понятию «ароморфоз», тогда как постепенные адаптивные преобразования при увеличении глубины обитания не выходили за рамки частных приспособлений.
Генеральная схема заселения океанических биотопов представляется сейчас в следующем виде (рис. 44). Шельфовые предки дали начало мезобентальным рыбам, а от них последовательно произошли бати- и абиссобентальные и хадальные формы. В этом процессе очень существенная роль принадлежала вытеснению на большие глубины примитивных представителей тех или иных таксонов, не выдержавших конкуренции с более прогрессивными формами. Заселение разных горизонтов пелагиали происходило от придонных и неритических предков независимо на разных глубинах и шло главным образом по пути прогрессивной ароморфной эволюции; в формировании батипелагической фауны существенное значение имело и заселение сверху, из мезопелагиали. Картина современного биотопического распределения океанических рыб осложняется вторичным подъемом представителей некоторых групп в более поверхностные горизонты, вторичным же возвращением из открытой пелагиали к склонам.
Глава 6. Современное состояние и перспективы океанического рыболовства
В начале этой книги я упомянул, что к числу объектов промысла относятся сотни видов рыб. «Тысячи, — поправил Н. П. Новиков, мой давний приятель, а ныне директор АзЧерНИРО — рыбохозяйственного института в Керчи,— ведь в "Словаре названий морских промысловых рыб" Г. У. Линдберга и соавторов значится три с лишним тысячи таких видов». Я задумался. Что же такое «промысловые рыбы» и не слишком ли расширенно мы толкуем это понятие? В упомянутом «Словаре», как сказано в предисловии к нему, «приводятся по возможности все виды рыб, служащие объектами рыболовства, а также обычные в уловах и имеющие народные названия». Это определение не отличается строгостью и к тому же представляется неправильным по своему существу — ведь многие рыболовные орудия облавливают водоемы безвыборочно, а народные названия есть почти у всех рыб, распространенных в пределах территории, которую этот народ занимает. Кстати говоря, несъедобных рыб практически не существует (кого только не приходилось дегустировать в экспедициях!), хотя некоторые из них могут употребляться в пищу только приготовленными соответственным образом. Я думаю, что в наше время «промысловыми рыбами» следует называть только тех, которые образуют локальные концентрации, достаточные для экономически оправданного облова, а отнесение к их числу рыб, случайно попадающих в снасти (так называемого «прилова»), неправомочно. К «потенциально промысловым рыбам» принадлежат такие виды, которые имеют достаточно большую биомассу, но не могут рентабельно облавливаться при современном состоянии промышленного рыболовства.
Промысловые и потенциально промысловые рыбы отличаются от не промысловых прежде всего высокой численностью и большей плотностью популяций. Отсюда следует, что они относятся к числу доминирующих видов своих ихтиоценов. Исключение составляют крупные хищники, занимающие в трофических пирамидах самые верхние уровни: их промысловое значение определяется не количеством, а качеством — очень высокой ценностью получаемого сырья. Доминирующие виды считаются оптимально приспособленными к конкретным условиям сообществ, хотя в чем именно проявляется эта приспособленность — сказать трудно. К примеру, из нерито-пелагических рыб (почти все они выходят и в открытые воды) преобладающее большинство — близкие подвиды или виды из родов Clupea (сельди), Sardinops (сардины иваси), Engraulis (анчоусы), Micromesistius (путассу), Trachurus (ставриды), Scomber (скумбрии) — имеют промысловое значение во всех районах, где они встречаются. Это правило не распространяется, однако, на род Theragra (минтай). Тихоокеанский минтай (Т. chalcogramma) принадлежит к числу важнейших промысловых рыб (в последние годы он твердо удерживает первое место в мире по вылову), а почти неотличимый от него атлантический вид (Т. finmarchica) остается известным по 4 коллекционным экземплярам, пойманным в юго-западной части Баренцева моря*.
* Т. С. Расс предполагает, что атлантического вида минтая не существует, а относимые к нему особи представляют собой тихоокеанских рыб, достигших берегов Норвегии «северным морским путем» в теплый год.
Известно также, что виды рыб могут с легкостью переходить из разряда непромысловых в категорию промысловых и обратно. Такие переходы могут быть следствием естественных флюктуации численности («волн жизни»), антропогенного воздействия, в том числе перелова, или природных катаклизмов. Хорошо известным примером такого катаклизма служит почти полное вымирание в 1882 г. хамелеоноголового хохлача (Lopholatilus chamaeleoniceps) (сем. малакантовые), обитающего у атлантического побережья США в водах верхней части континентального склона (глубина 100 — 300 м). Хохлач живет в очень узком диапазоне температуры (8 — 12°), так что даже относительно небольшое охлаждение воды привело тогда к его массовой гибели (по приближенной оценке на поверхности было насчитано около 1,5 млрд мертвых рыб) и вид надолго выбыл из списка промысловых объектов. Случаи массовой смерти придонных рыб наблюдались и в других районах океана.