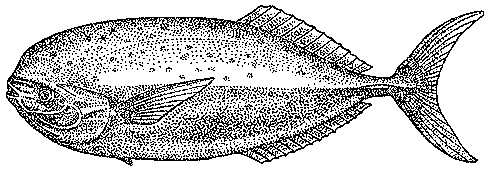
На мой взгляд, океанический двузуб самим фактом своего существования представляет одно из наиболее наглядных доказательств продолжающегося заселения эпипелагиали прибрежными рыбами. Это — пример недавнего вселенца, нашедшего в новом биотопе благоприятные условия, но еще не успевшего приобрести специфических адаптации к нему. На противоположном полюсе я поместил бы луваря (Luvarus imperialis) (рис. 43) — удивительную, внешне ни на кого не похожую рыбу, которую в продолжение многих лет считали родственной скумбриевым или ставридовым. Новейшие анатомические исследования показали, что луварь ведет начало от прибрежных рыб-хирургов, но длительная эволюция в условиях открытого океана сделала его совсем непохожим на своих предков.
Рис. 43. Луварь — рыба, родственные связи которой долго оставались непонятными
В эпипелагиали явно преобладают филогенетически молодые группы костистых рыб (в частности, окунеобразные). Основываясь на этом факте, я предполагал ранее, что до появления этих вселенцев верхние слои открытого океана были заселены низшими (мягкоперыми) рыбами, которые, не выдержав конкуренции, оказались вытесненными вниз и дали начало современному населению средне-глубинной зоны. Сейчас я считаю это умозрительное предположение крайне маловероятным. Существует гораздо больше оснований для иной точки зрения: главным источником формирования мезопелагического ихтиоцена послужили придонные и придонно-пелагические формы, обитавшие на материковом склоне.
Представление о происхождении мезопелагических рыб от мезобентальных и мезобентопелагических предков основывается на следующих соображениях. Специфические подотряды и семейства мезопелагических рыб относятся к таким отрядам (угреобразные, аулопообразные, миктофообразные, трескообразные, бериксообразные и др.), основная масса представителей которых обитает на континентальном склоне, и являются наиболее специализированными таксонами этих групп. Семейство Chiasmodontidae наиболее родственно верхнемезобентальному сем. Champsodontidae. Мезопелагические роды в семействах, распространенных и вне рассматриваемого биотопа, принадлежат к группам, характерным в основном для мезобентали (Isistius и Euprotomicrus — к сем. колючих акул, Xenophthalmichthys — к сем. серебрянковых, Scopelengys — к сем. неоскопеловых, Brotulotaenia — к сем. ошибневых, Cynomacrurus и Mesobius — к сем. долгохвостовых, Diretmus — к сем. диретмовых, Diplospinus и Nealotus — к сем. гемпиловых, Ectreposebastes — к сем. скорпеновых, Nectoliparis и Rhodichthys — к сем. морских слизней). В то же время мезопелагические виды в преимущественно придонных родах, по-видимому, очень редки: единственный известный мне пример представляет Nezumia parini,— описанный К. Хаббсом и Т. Ивамото пелагический долгохвост из восточной тропической Пацифики.
Бати- и абиссопелагический ихтиоцены в основном формировались, по-видимому, тем же путем, что и мезопела-гический. Судя по распространению рецентных родственных групп, специфические высокоспециализированные глубоководные семейства угреобразных (пилосошниковые, циемовые, мешкоротовые) и удильщикообразных (11 семейств подотряда Ceratioidei) ведут свое начало от донно-придонных рыб нижней части материкового склона. В пелагиали больших глубин значительная роль принадлежит и представителям таких семейств, которые еще не полностью утратили связь с дном — гладкоголовам и платитроктовым, откладывающим донную икру, а также долгохвостам, афионидам и др. В то же время наличие в составе глубинной пелагической ихтиофауны родов, принадлежащих к преимущественно мезопелагическим семействам гоностомовых, малакостеевых и миктофовых, указывает на частичное заселение этой зоны из средних горизонтов.
Нужно сказать, что представители отдельных групп мезопелагических рыб, принадлежащих к макропланктону, оказались способными выйти за пределы исходного биотопа и значительно расширить адаптивную зону обитания этих групп. Наиболее характерные примеры внедрения во все смежные участки среды дают светящиеся анчоусы (сем. миктофовые) которые справедливо считаются одной из самых процветающих групп мезопелагического ихтиоцена. Тем не менее около трети видов семейства (а их число достигает 220) временно или постоянно входит в состав других сообществ. Так, около 40 видов поднимаются ночью в эпипелагиаль (по меньшей мере один из них — Benthosema pterotum, наблюдавшийся в восточной тропической Пацифике у поверхности даже в светлое время суток, может образовывать там жилую, т. е. не мигрирующую, популяцию), около 10 видов живут в батипелагиали и около 20 — вторично возвратились в бентопелагические биотопы (туда, где когда-то обитали предковые формы миктофид) и в некоторых случаях достигли там высокой численности. Вторичнобентопелагические виды возникли и в других мезопелагических семействах — среди топориковых (достаточно вспомнить мавролика), фотихтовых, гоностомовых, меланостомиевых, меламфаевых. Такие виды есть также среди мезобатипелагических, в основной своей массе глубоководных удильщиков церациоидей: это представители родов Diceratias и Phrynichthys. Мало того, один из видов сем. тауматихтовых — Thaumatichthys axeli является, по всей вероятности, вторично-донной рыбой (единственный в своем роде случай «бентизации», контрастирующий с обычной «пелагизацией» генеративно донных форм).
Говоря о заселении рыбами океанических биотопов, нельзя обойти вниманием вопрос о параллелизмах и конвергенциях в их эволюции, сопровождавших освоение тех или иных участков среды обитания. Этот вопрос, издавна привлекавший внимание исследователей, был в недавнее время детально разобран Н. Маршаллом, что позволяет ограничиться здесь рассмотрением отдельных, наиболее ярких проявлений функционального и структурного сходства систем и органов у рыб, входящих в состав разных сообществ. В эпипелагиали наиболее поражает разительное сходство между акулами ламнидами (к ним относятся, напомню, сельдевые акулы, мако и кархародон) и тунцами. Это касается не только общей формы идеально обтекаемого торпедообразного тела, его окраски, положения и относительных размеров непарных плавников, наличия стабилизирующих килей на чрезвычайно узком хвостовом стебле, несущем мощный хвостовой плавник в форме полумесяца. Еще более замечательны конвергенции в строении внутренних органов, и главная из них несомненно состоит в необычайном развитии подкожной сосудистой системы, питающей туловищную мускулатуру и обеспечивающей «теплокровность» у этих совсем не родственных рыб. И у тунцов и у ламповых акул мощно развиты наиболее активные красные мышцы латеральной мускульной системы. Волокна этих мышц окружены «чудесной сетью» капилляров, которая обеспечивает интенсивный теплообмен между артериальной кровью, исходно (в жабрах) имеющей температуру морской воды, и венозной кровью, нагретой за счет мышечной работы. В результате этого обмена температура тела тунцов и акул оказывается на 6 — 10° выше «забортной» температуры, и это приводит к значительным энергетическим выгодам (ведь скорость всех ферментативных реакций повышается в таком случае в 2 — 3 раза). Крупные тунцы (виды рода Thunnus) и акулы сем. Lamnidae могут считаться «идеальными нектерами» — как по «крейсерским» достоинствам, так и по максимальной скорости на спринтерской дистанции они не имеют соперников ни среди других рыб (исключение составляют мечевидные, достигшие тех же результатов), ни среди кальмаров и морских млекопитающих.
Примеры конвергенции и параллелизмов среди глубоководных пелагических рыб очень многочисленны, и их полное перечисление заняло бы здесь слишком много места. Нельзя не упомянуть тем не менее бросающегося в глаза сходства в окраске тела — серебристого у многих верхнемезопелагических видов (из батилаговых, топориковых, миктофовых, палочкохвостых и др.) и черного или коричневого у всех форм, обитающих на глубине более 1000 м. Органы свечения независимо возникли у колючих акул (отряд катранообразные) и у костистых рыб, принадлежащих к отрядам лососеобразных, стомиеобразных, аулопообразных, миктофообразных, окунеобразных (род Pseudoscopelus). Телескопические глаза характерны для представителей отрядов лососеобразных (Xenophthalmichthys, Opisthoproctus), аулопообразных (Gigantura, Scopelarchus, Evermannella), миктофообразных (Protomyctophum), опахообразных (Stylephorus). Многие стомиеобразные и все удильщики приманивают добычу своеобразными удочками со светящейся приманкой. Для всех хищников характерны приспособления для захвата крупной добычи — огромные пасти, зубатые челюсти, редуцированные жаберные тычинки, сильно растяжимые желудки и стенки тела. У идиакантов и удильщиков карликовые самцы значительно мельче самок. Параллельным путем возникли и вертикальные миграции мезопелагических рыб.