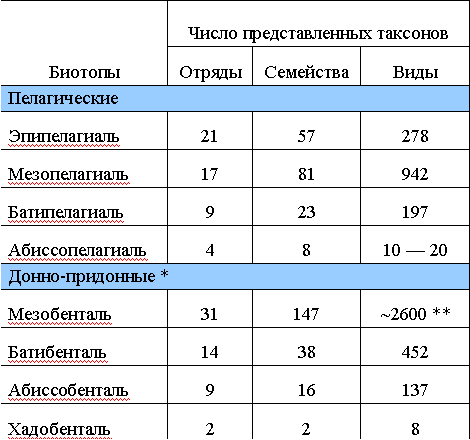
Таблица 5
Систематический состав (число семейств) морских прибрежных и океанических ихтиоценов
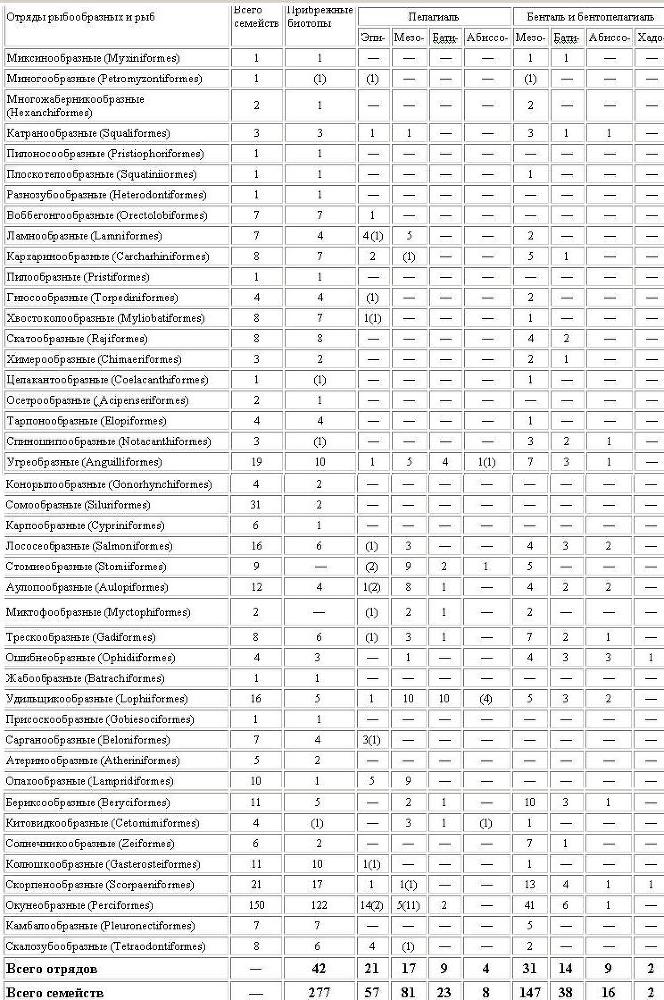
В целом ихтиофауна открытого океана образует значительную но объему и хорошо обособленную часть мировой фауны рыб (табл. 6). Она включает, по самым предварительным подсчетам (для более точных оценок время еще не наступило), около 1100 видов, живущих в толще воды без какой-либо связи с дном и около 2800 видов бентопелагического комплекса (без талассосублиторальных видов, число которых, по самой грубой прикидке, составляет не менее 300 — 400), в том числе почти 1700 видов, не выходящих на шельф. В полном объеме океаническая ихтиофауна содержит, таким образом, около 3900 (2200 без шельфо-батиальных форм) видов (см. табл. 2 — 4), т. е. составляет более четверти всех морских рыб (прибрежных рыб насчитывается около 9,5 тыс. видов). Несколько отличающиеся оценочные цифры приводит Д. Коэн, по подсчету которого число видов прибрежных рыб достигает 9100, а океанических — 2500 (в том числе 255 эпипелагических, 1280 глубоководных донно-придонных и 1010 глубоководных пелагических).
Таксономическая обособленность отдельных биотопических группировок океанических рыб довольно сильно различается. Для эпипелагиали, например, специфичны всего 16 семейств рыб (30% от числа представленных в биотопе), причем большая их часть характерна для ее нижних слоев, пограничных с мезопелагиалыо. Полностью океаническим является, строго говоря, только одно из верхнеэпипелагических семейств — макрелещуковые. Специфичность на родовом и видовом уровнях достигает тем не менее 50% списочного состава.
Глубоководные пелагические ихтиоцены значительно более своеобразны. Их самобытный облик определяется главным образом наличием ряда характерных таксонов высокого ранга, таких, как отряды стомиеобразных, миктофообразных, китовидкообразных, подотряд глубоководных удильщиков и др., очень разнообразных в видовом отношении. Все они отсутствуют в прибрежной зоне и очень слабо представлены в толще воды над материковым склоном. Половина мезобатипелагических семейств (42 из 83), большинство родов (61%) и почти все виды (85%) также специфичны для этих биотопов.
Таблица 6
Систематическое разнообразие океанических ихтиоценов
(число представленных таксонов)
* Весь бентопелагический комплекс (бенталь + бентопелагиаль).
** Без эпимезобентальных около 1500 видов.
Придонные ихтиоцены открытого океана представляются менее обособленными, но и они имеют характерные отряды (целакантообразные, спиношипообразные) и подотряды (например, гладкоголововидные). Число специфических семейств относительно невелико — всего 20% (27 из 137), хотя на уровне родов и видов эти таксоцены вполне четко отличаются от прибрежных и пелагических, а также друг от друга.
В целом наиболее резко выражены границы между совокупностями пелагических и донных таксоценов рыб, причем степень выраженности этих границ явно увеличивается с глубиной. В вертикальной последовательности бентальных и бентопелагических ихтиоценов самая четкая разделительная полоса соответствует краю континентальной отмели, но, как указывалось в предыдущей главе, многие преимущественно шельфовые виды все же опускаются в верхние участки материкового склона. Дальнейшие изменения таксономического состава — от мезобентали к батибентали и далее к абиссобентали и хадали — происходят постепенно и наиболее заметны на видовом уровне. В толще воды границы между эпи-, мезо- и батипелагиалью, несмотря на существование зон смешения (экотонов), выражены более четко, чем границы между гомологичными донно-придонными биотопами.
Исходным материалом для формирования рецентной фауны океанических рыб бесспорно были шельфовые и неритические предковые формы. Из этого постулата со всей определенностью следует, что именно прибрежные участки (или по меньшей мере некоторые из них) действовали в определенные периоды времени, а быть может, действуют и сейчас как эволюционные центры, продуцирующие виды, способные к заселению новых биотопов за пределами этих участков, т. е. к переходу в новую среду обитания*. Очевидно также, что прямо из шельфо-неритической зоны могли заселяться только непосредственно прилегающие к ней биотопы, которые, в свою очередь, обеспечивали заселение контактирующих с ними участков среды. Последовательность появления вселенцев в том или ином биотопе может в первом приближении определяться их ранжированием по систематической обособленности: наличие видов, общих с исходным (донорским) биотопом, говорит о совсем недавнем или еще не закончившемся вселении, тогда как дивергенция на видовом и родовом уровне, а тем более на уровне семейств, подотрядов и отрядов свидетельствует о все увеличивающемся сроке, прошедшем со времени успешной колонизации.
* Альтернативой такому представлению могло бы служить только явно абсурдное в данном аспекте предположение о некогда сплошном распространении предковых форм в относительно однородной среде и последующем расчленении их ареалов в результате образования физических барьеров (так называемая «викариация» в понимании сторонников кладизма).
Основываясь на этих представлениях, рассмотрим теперь, как происходило заселение донно-придонных океанических биотопов. В мезобентали очень обычны рыбы, характерные и для шельфовой зоны (см. табл. 4). Большинство этих эврибатных эпимезопелагических видов входит в состав экотонного сообщества кромки шельфа и самой верхней батиали и не опускается глубже 400 — 500 м. Среди них, несомненно, есть и такие формы, которые потенциально способны к адаптивной модификации, дающей возможность закрепиться на глубинах материкового склона.
Примеры такого закрепления показывают мезобентальные виды преимущественно шельфовых родов, например Saurida kaianus (сем. Synodontidae) или Chaetodon guyotensis (Chaetodontidae). Есть в этом биотопе и специфические роды, принадлежащие к преимущественно шельфовым семействам — Ostichthys (сем. Holocentridae), Aphanopus и Assurger (Trichiuridae), Bembradium (Platycephalidae) и др. К эндемичным мезобентальным семействам относятся плащеносные акулы и акулы-домовые, ложноромбовые и шестижаберные скаты из щележаберных, глубоководные солнечники, хаунаксовые и холлардиевые из костистых рыб. Только в мезобентали сохранился до нашего времени: и единственный представитель кистеперых рыб — латимерия.
В глубине души я всегда надеялся увидеть латимерию живьем, но мне так и не довелось, к превеликому сожалению, присутствовать при ее поимке. Ближе всего к живому целаканту я был, пожалуй, в апреле 1983 г., когда судно «Рифт» проводило траления на глубине 400 — 500 м у северо-западного побережья Мадагаскара, всего в какой-нибудь сотне миль от Коморских островов. Увы, безрезультатно... Правда, с фиксированным экземпляром мне познакомиться все-таки довелось. Дело в том, что еще в 1972 г. наш институт приобрел по специальному указанию директора – члена-корреспондента Академии наук А. С. Монина, одну латимерию для предполагавшейся выставочной коллекции. Покупка была оплачена через Париж (острова еще были колонией Франции), но для получения экспоната «Дмитрий Менделеев», работавший тогда в Индийском океане, был послан прямо на остров Гранд-Комор. Вот как описывает этот эпизод участник экспедиции и-мой хороший друг Виктор Нейман: «Разрешения от местных властей на посещение Коморских островов мы в тот раз не получили и должны были принять рыбу на борт на внешнем рейде... После довольно долгого томительного ожидания к судну подошел, наконец, старенький катер, на палубе которого в открытом жестяном ящике находилась с нетерпением ожидавшаяся всеми диковина. Вид ее нас, однако, разочаровал до крайности — грязно-рыжая рыбина, по-видимому, давным-давно ждавшая нас в растворе формалина, отнюдь не блистала красотой». Действительно, целакант прибыл в Москву в ужасном состоянии – его полутораметровое тело было почти полностью покрыто плотной, как панцирь, коркой красноватой ржавчины, удалить которую механическим путем, не повредив при этом внешних покровов, было невозможно. Очистку латимерии по предложению сотрудника нашей лаборатории Дмитрия Астахова, по праву заслужившего звание «лучшего химика среди ихтиологов», вели с применением растворителя окиси железа — так называемого трилона Б. С результатами этой «реставрационной деятельности» можно ознакомиться в фойе Института океанологии им. П. П. Ширшова (Москва, ул. Красикова, 23), где обповлениый целакант в гордом одиночестве выставлен для обозрения в большом плексигласовом сосуде, доверху заполненном фиксатором. Жаль только, что рыба навсегда утратила свою сиренево-синюю окраску и стала теперь тускло-серой, но такова уж судьба всех музейных экспонатов.