Таким образом, для эпипелагических рыб типичны слабо выраженная избирательность питания и как ее следствие наличие широких пищевых спектров. Большинство эпипелагических рыб питается практически любым доступным кормом, подходящим по размерам и имеющимся в данном месте в достаточном количестве. В схеме трофической структуры океанской пелагиали (рис. 16) рыбы занимают несколько верхних уровней — до пяти в тропических водах, характеризующихся наибольшей сложностью пищевых взаимоотношений.
Распространение. Хорошо известно, что границы ареалов эпипелагических рыб, принадлежащих к нектону (т. е. способных к активным горизонтальным перемещениям), определяются главным образом температурой воды. Связь распространения с температурными условиями показана для многих рыб — акул, лососей, летучих рыб, тунцов, марлинов и др. Двукрылая летучая рыба Exocoetus volitans, например, встречается при температуре 22 — 29°, будучи обычной при 24 — 28°. В результате область распространения этого вида в наиболее теплой западно-экваториальной части Тихого океана была прервана осенью 1961 г. примерно на 15° по широте (т. е. на 900 морских миль), а в центральной части океана, где в области экватора температура несколько понижена благодаря поднятию глубинных вод, не имела такого разрыва. Минимальные значения температуры, отмеченные для разных видов массовых скумбриевидных рыб открытого океана в период нагула, варьируют от 10 — 12° у длинноперого тунца и меч-рыбы до 18 — 20° у желтоперого тунца и синего марлина. Температурный диапазон встречаемости в океане лососей рода Oncorhynchus полностью укладывается в пределы 2,3 — 12°. причем наиболее холодолюбивым видом является нерка, обычная при 2,3 — 9°, а наиболее теплолюбивым — кижуч, предпочитающий температуру 7 — 12°. Подобные примеры, количество которых может быть многократно увеличено, вполне убедительно демонстрируют значение температуры воды как главного фактора, лимитирующего географическое распространение нектонных рыб.
Этот фактор нельзя, конечно, считать единственным. Ограничивающая роль может принадлежать и другим показателям, например содержанию в воде кислорода (2 мл/л — предельное значение для опускающегося вглубь альбакора), солености воды (35,5%о — минимальная, как полагают, величина для приближающейся к берегам длиннокрылой акулы) или количественному распределению кормовых организмов.
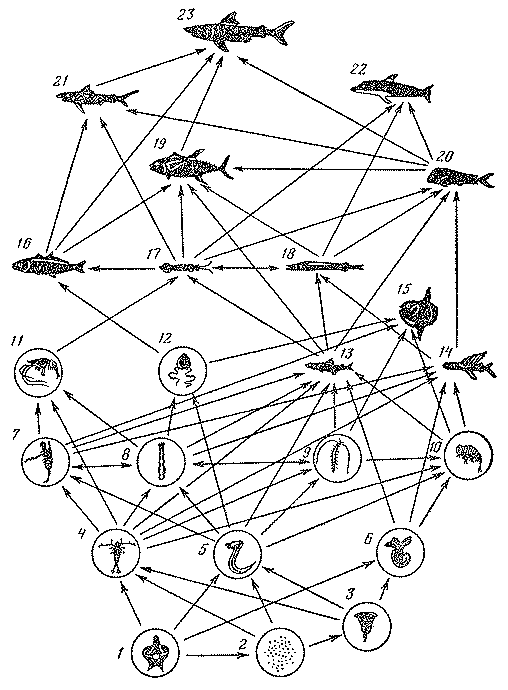
Рис. 16. Схема основных пищевых связей в сообществах тропической зоны
1 — фитопланктон; 2 — бактерии; 3 - 12 — зоопланктон; 13 — миктофиды; 14 — летучие рыбы; 15 — рыбы-луны; 16 — строматеевидные; 17 — кальмары; 18 — гемпиловые; 19 — тунцы; 20 — корифены; 21 — синяя акула; 22 — дельфины; 23 — кархародон
В ареалах нектонных рыб, как уже говорилось, можно различить три структурные части — область размножения, нагульную область и область выселения планктонных стадий раннего развития. Пространственное взаиморасположение репродуктивной и нагульной частей ареала у разных видов может существенно варьировать. В некоторых случаях они полностью совпадают, в других — частично перекрываются, в третьих — нацело разобщены.
Правда, при современной изученности распространения нектонных рыб невозможно выделить структурные части ареалов для большинства эпипелагических видов. Достаточно детальные представления имеются лишь о распространении относительно немногих рыб. Ареал северотихоокеанской сайры, например, состоит из репродуктивной части, занимающей южные районы области распространения (на периферии Северного субтропического круговорота), нагульной части, приходящейся на субарктические воды, и области выселения мальков в заливе Аляска. Полная или частичная пространственная разобщенность нерестовой и нагульной частей ареала имеется также у сельдевой и синей акул, скумбрещуки, субтропической летучей рыбы Hirundichthys rondeletii, длинноперого тунца, полосатого марлина, меч-рыбы и некоторых других видов. При этом основные структурные части ареалов некоторых широко распространенных нектонных рыб эпипелагиали могут располагаться в значительно различающихся условиях среды.
Значительные перемещения многих нектонных рыб в пределах своих ареалов, связанные с периодическими нерестовыми, нагульными и зимовальными миграциями, приводят к существенным изменениям границ распространения отдельных видов в разные сезоны. Особенно сильно меняется распространение в океане у тех рыб, которые обитают в водах, подверженных заметным сезонным изменениям температуры,— у сайры, лососей, некоторых тунцов и акул и т. п.
Число планктонных рыб в эпипелагиали невелико: если исключить из рассмотрения никтоэпипелагических мигрантов, к ним относятся лишь карликовые виды семейства макрелещуковых, а также пассивные в течение всей жизни гиганты — рыбы-луны (по всей вероятности, сюда следует добавить сельдяных королей, вогмеров, парусных морских лещей рода Pteraclis). Можно думать, что географическое распространение этих рыб определяется, как и распространение других планктонных животных, циркуляцией вод океана, а основы их ареалов более или менее соответствуют крупномасштабным системам замкнутых или полузамкнутых круговоротов.
Миграции. Характерной чертой многих эпипелагических рыб служит их способность к направленным перемещениям в пространстве — вертикальным и горизонтальным миграциям. Оставляя в стороне миграции никтоэпипелагических видов (они будут рассмотрены в гл. III), следует отметить, что наибольшие по размаху вертикальные перемещения совершают тунцы рода Thunnus и особенно меч-рыба. По имеющимся данным, желтоперый тунец может опускаться до глубины 160 м, большеглазый — до 250 м, а длинноперый еще глубже — до 380 и даже 600 м. Меч-рыба также была однажды сфотографирована на глубине, превышающей 600 м.
Активные горизонтальные миграции некоторых эпипелагических рыб, обусловленные пространственным разобщением районов нереста и нагула в соответствии с различиями абиотических (прежде всего температурных) и биотических (в основном кормовых) условий, достигают очень большой протяженности и часто имеют довольно сложный характер. Они еще недостаточно изучены и лишь в немногих случаях подтверждены данными мечения, позволяющими прослеживать их пути. Иногда, впрочем, такие миграции хорошо заметны по перемещениям массовых скоплений и районов промысла. У океанических скумбриевидных рыб они достигают наибольшего размаха: у них отмечены даже сверхдальние трансокеанические миграции.
Особенно значительные перемещения совершают, по-видимому, генеративно тепловодные (т. е. имеющие тропическое происхождение) рыбы, выходящие для нагула за пределы тропических вод. Так, детальное изучение биологии длинноперого тунца (альбакора) в северной части Тихого океана показало, что для этого вида в период, предшествующий наступлению половой зрелости, характерны весьма сложные перемещения.
Опыты по мечению длинноперого тунца начались в 1952 г., а уже на следующий год было получено неоспоримое доказательство трансокеанских миграций: рыба, помеченная в районе Калифорнии в августе 1952 г., была обнаружена в японских водах через 323 дня после выпуска. В последующие годы в западной части Тихого океана было поймано еще пять рыб, кроме того, два тунца были возвращены из центральной части океана (район о-ва Мидуэй). Средняя скорость перемещения, по данным мечения, достигает 16 — 17 миль (около 30 км) в сутки. Мечение длинноперого тунца производилось также в центральной части Тихого океана, восточнее Японии. Рыбы, помеченные в центре океана, ловились впоследствии и у Японии, и у Калифорнии, а тунцы, выпущенные в японских водах, добывались лишь в средней части океана.
Результаты мечения, данные по размерному и возрастному составам уловов, а также сведения о сезонных перемещениях районов промысла позволили представить в общем виде картину миграций неполовозрелого альбакора в северной половине Тихого океана. В промысловых уловах у Японии и Калифорнии длинноперый тунец, как правило, встречается, начиная с двухлетнего возраста, причем в восточной части океана добываются рыбы в возрасте от 2 до 5 лет, а в западной — от 1 до 10 лет. Исходя из возможности одной трансокеанской миграции в год, американские ученые предложили схему миграций этого вида, которая вполне согласуется с имеющимся фактическим материалом. Эта логичная картина, воссозданная в начале 60-х годов, рассматривается теперь лишь как одна из возможных моделей миграционного цикла альбакора.