Пока эмбрион состоял из нескольких клеток, каждая из них занимала достаточно большую долю в общем объеме. Часть поверхности каждой клетки была обращена в окружающую среду. Когда в ходе дробления получается тридцать две или шестьдесят четыре клетки, уже достаточно мелкие по сравнению с размерами сферического зародыша, некоторые из них оказываются окружены со всех сторон другими клетками. Другие все еще имеют контакт с внешней средой – примерно одна шестая их мембраны обращена наружу. Клетки «чувствуют», окружены ли они другими клетками или какая-то их часть соприкасается только с жидкостью. Эту информацию они используют для того, чтобы определиться с дальнейшими действиями. Клетки, имеющие «свободную поверхность», активируют ряд ранее неактивных генов и образуют первую ткань эмбриона – трофэктодерму. Клетки, окруженные другими клетками, эти гены не активируют. Использование в качестве источника информации простого физического признака – наличия свободной поверхности – избавляет эмбрион от необходимости иметь заранее подготовленный пространственный план. Кроме того, клеткам не нужно «знать» свое точное расположение в эмбрионе. Им нужно только определить, есть у них свободные участки поверхности или нет.
Задача трофоэктодермы заключается только в создании структур для структурной поддержки и питания эмбриона. Частью ребенка она никогда не станет.[22] В первую очередь трофэктодерма перекачивает в эмбрион жидкость. Эта жидкость накапливается, что приводит к образованию обширной полости (рис. 6). Формирование этой полости, в свою очередь, приводит к тому, что внутренние клетки группируются на внутренней поверхности трофэктодермы в виде эксцентрично расположенного комка, который называется «внутренняя клеточная масса». Если судить только по внешнему виду, эти клетки кажутся менее интересными, чем активные клетки трофэктодермы, постоянно занятые питанием, накачкой жидкости, внедрением в материнский организм. Однако именно из внутренней клеточной массы и формируется ребенок. Иногда эта масса делится на два комочка, каждый из которых развивается, формируя в итоге человеческий организм. Такие близнецы, как и те, о которых рассказывалось в главе 2, будут генетически идентичны, но на этот раз они будут расти в одной общей трофэктодерме. В то же время у каждого из близнецов будет свой желточный мешок и амниотическая полость (см. далее). Таким образом, они надежно изолированы друг от друга. Это самый распространенный механизм появления однояйцевых близнецов, на его долю приходится две трети всех случаев. (Третий, крайне редкий вариант появления близнецов будет описан несколько позже.)
К тому времени, когда клетки трофэктодермы закачают внутрь эмбриона объем жидкости, достаточный для создания полости, и создают вздувшуюся структуру, эмбрион должен покинуть фаллопиеву трубу матери, где произошло оплодотворение, и переместиться в матку. Этот орган в норме как бы немного «сдут»: его стенки слипаются, как снятая с руки резиновая перчатка. Особенно «сдувшейся» матка выглядит через неделю после овуляции. Поэтому весьма вероятно, что ранний эмбрион, попав в матку, вскоре столкнется с ее внутренней поверхностью. Когда это произойдет, он закрепляется на ней, используя специальный набор адгезивных молекул. После прикрепления клетки эмбриона образуют новые белки, которые позволяют им внедриться между клетками стенки матки.[23] В течение нескольких часов «пальчики» клеток эмбриона проникнут в ткани матери, и образуется плацента. Многие клетки матки при этом разрушаются, а их остатки становятся пищей для эмбриона. Реакция матери на эту атаку приводит к дальнейшему отмиранию ткани. Таким образом, через десять дней после оплодотворения формируется полость, в которой легко умещается эмбрион. У людей и некоторых животных внутренняя выстилка матки обрастает эмбрион, скрывая место имплантации.
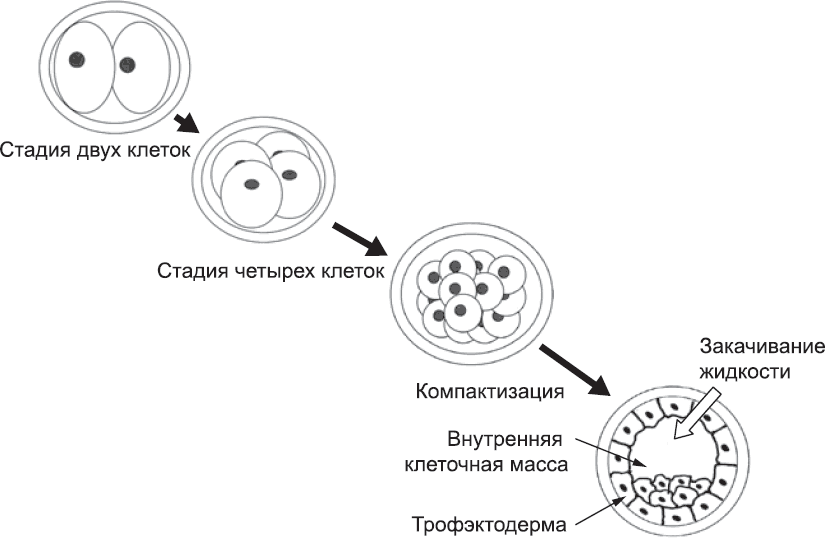
Рис. 6. Схема перехода от стадии двух клеток к стадии образования полости и затем к стадии обособления трофэктодермы и внутренней клеточной массы. Эмбрион заключен в плотную желеобразную «капсулу», прежде служившую оболочкой яйцеклетки; она носит название вителлинового слоя, или zona pellucida
Образ жизни эмбриона человека близок к паразитическому, но не стоит принимать эту метафору слишком близко к сердцу. В любом случае это единственный способ вынашивания потомства, а значит, выживания у нашего вида. Мать не просто терпит паразитизм ребенка, а поощряет его за счет постоянного взаимодействия. Если построенный на сигнальных молекулах «диалог» между маткой и эмбрионом прекращается, эмбрион не может оставаться имплантированным, и беременность прерывается.[24]
Иногда эмбрион движется по женским половым путям от места оплодотворения в маточной (фаллопиевой) трубе чересчур медленно и не достигает матки к тому времени, когда уже готов к имплантации. Одной из распространенных причин этого медленного перемещения является повреждение выстилки маточной трубы бактериями Chlamydia.[25] В наше время это часто встречается у молодых женщин.[26] Эмбрион, достигший определенной стадии развития, пытается имплантироваться там, где он находится в данный момент, даже в яйцеводе. Это ведет к внематочной беременности. Маточная труба не приспособлена для поддержания роста плода ни физически (по размеру и растяжимости), ни физиологически (в плане возможности обеспечить питание и приток крови). Во многих случаях это приводит к выкидышу. Иногда выкидыш приходится вызывать искусственно для спасения жизни матери.
До начала специализации клетки внутренней клеточной массы могут дать начало любой клетке организма. Во всяком случае, так происходит у мышей: на людях подобные эксперименты не могут проводиться по этическим соображениям. Для описания спектра типов клеток, в которые может дифференцироваться та или иная клетка эмбриона, исследователи часто используют специальную древовидную диаграмму, отображающую типы клеток и последовательность дифференцировки. Пример такой диаграммы приведен на рис. 7. Все возможные варианты в конечном счете происходят от одного типа клеток, который на такой диаграмме представляет собой ствол дерева. Поэтому эти клетки называют стволовыми клетками. (Это буквальный перевод немецкого термина «Stammzelle», который предложил в 1909 г. Александр Максимов, изучавший «древо» клеток крови.) В зависимости от конкретной схемы термин «стволовые клетки» может быть применен как к клетке, которая может создать только несколько различных типов клеток организма, так и к клеткам с большими потенциями.
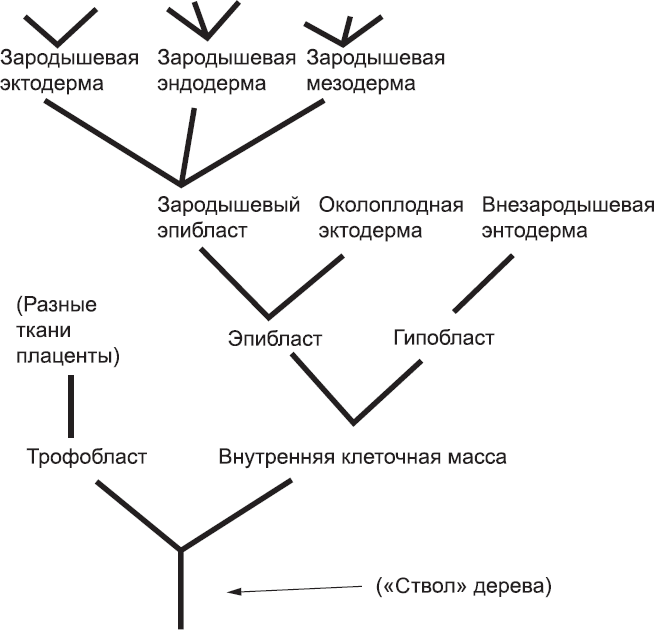
Рис. 7. Типичная древовидная диаграмма, которая обобщает данные о траекториях клеточных дифференцировок – клетки ранних стадий развития дают начало клеткам более поздних стадий (или дифференцируются в них). Эта диаграмма, которую следует читать снизу вверх, отражает события, описанные в этой и последующих главах. Древо всего процесса развития человека было бы огромным и содержало бы сотни ветвей. Нижняя точка таких диаграмм всегда принимается за «стволовую клетку» независимо от того, находится ли эта клетка на самом раннем этапе развития или уже прошла определенное расстояние «вверх» вдоль ствола дерева. Об использовании термина «стволовые клетки» будет рассказано позже
В последнее время понятие «стволовые клетки» несколько сузилось. Многие авторы утверждают, что стволовыми клетками следует называть только клетки, способные поддерживать собственную популяцию, а также давать начало другим типам клеток, расположенным на диаграмме над ними. (Термин, предложенный Максимовым, в явном виде не предполагал подобной трактовки).[27] К счастью, эта терминологическая тонкость не влияет на применимость термина «стволовые клетки» к внутренней клеточной массе, которая показана на рис. 7 в нижнем правом углу, потому что ее клетки могут поддерживать свою популяцию, даже если удалить их из эмбриона и поместить в пробирку. Они дают начало всем тканям организма. Поэтому клетки внутренней клеточной массы иногда называют эмбриональными стволовыми клетками (ЭСК), особенно в тех случаях, когда их извлекают из эмбрионов и выращивают в культуре.[28]