У собак, как и у всех млекопитающих, очень сложная нервная система, но подобный условный рефлекс встречается и у более простых позвоночных. Например, тропическая рыба Chromobotia macrocanthus быстро научается ассоциировать бренчание банки с кормом с кормлением и при этом звуке впадает в характерное возбужденное состояние. Такое научение было показано даже у плодовых мушек, хотя их нервная система устроена значительно проще, чем у собак. Если обычных плодовых мушек поместить в трубку, проходящую между двумя камерами, в каждой из которых свой характерный запах, мушки перемещаются из камеры в камеру хаотично, без особых предпочтений. Если сначала «познакомить» их с одним из запахов, никакой разницы в последующем поведении не будет. Если же сначала «познакомить» их с одним из запахов, подвергая их при этом серии электрических разрядов, а затем поместить в трубку, мушки будут избегать «знакомого» запаха и переместятся в другую камеру. Очевидно, они «научились» ассоциировать запах с электрическими разрядами.[263],[264]
Хебб предположил, что принимающие клетки могут биохимически менять эффективность ответа на сигналы от конкретных синапсов в соответствии с правилом: эффективность ответа на сигналы от синапса увеличивается, если синапс передает импульс в то же время, когда принимающая клетка тоже передает импульс. Чтобы проиллюстрировать это, рассмотрим простую модель части нервной системы плодовых мушек, которые подвергались эксперименту с запахами и электрическим разрядом. В этой модели (рис. 72) задействовано четыре типа нейронов (подобное упрощение далеко от реальности, но поможет объяснить основной принцип). Нейроны типа O1 активируются в присутствии запаха 1, а нейроны типа O2 активируются в присутствии запаха 2. Нейроны типа E активируются под воздействием электрического тока, а нейрон R, к которому подключены все остальные нейроны, при активации вызовет у мушки реакцию «спасения бегством». До обучения связи нейронов обоих типов (O1 и O2) с нейроном R слишком слабые, чтобы активировать его, так что ни один запах не подталкивает мушку к бегству. Однако связь нейрона R с нейронами типа E достаточно сильная, чтобы активировать нейрон R при стимуляции электрическим током. Если при использовании электрического тока и активировании нейрона R с помощью сигналов нейронов типа Е присутствует запах 1, будут активны нейроны типа O1. Таким образом, условие Хебба будет выполнено, и реакция нейрона R на синапсы O1 усилится.
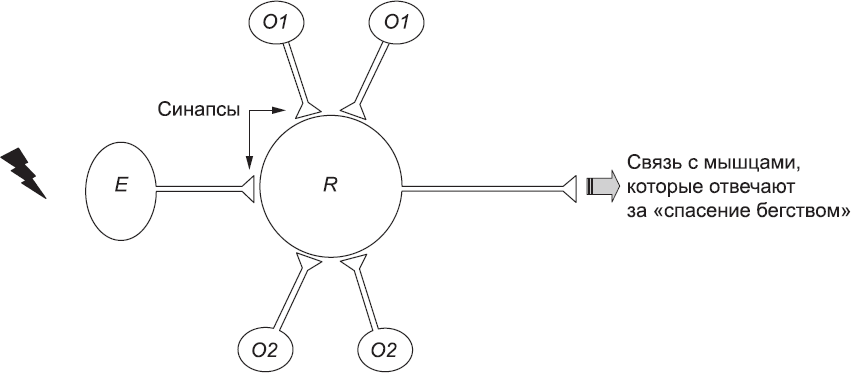
Рис. 72. Упрощенная модель выработки условного рефлекса у плодовых мушек. Сначала нейрон R получает слабые сигналы как от нейронов O1, реагирующих на запах 1, так и от нейронов O2, реагирующих на запах 2, а также сигналы от нейронов типа E, которые интенсивно стимулируются электрическим током. На самом деле в процессе участвует гораздо большее количество нейронов каждого типа, но на рисунке для ясности показаны лишь некоторые из них
Если это будет происходить достаточно часто, то реакция будет настолько сильной, что одного только импульса нейронов О1 будет достаточно, и плодовая мушка станет избегать запаха 1, не дожидаясь удара током. Реакция на нейроны O2 при этом не будет усиливаться, и запах 2 не будет вызывать у мушек желания «спастись бегством».
За десятилетия после публикации гипотезы Хебба были открыты некоторые биохимические механизмы, лежащие в основе описанного процесса. Во многих исследованных нервных системах, включая нервную систему плодовой мушки, в синапсах действует определенный нейромедиатор – глутамат. У принимающих клеток есть два типа рецепторов для этого нейромедиатора. Рецепторы типа AMPAR работают просто: каждая молекула AMPAR, связавшись с глутаматом, активирует комплекс белков внутри принимающей клетки и вносит свой скромный вклад в попытки вызвать передачу импульса. Если достаточное количество молекул AMPAR свяжется с глутаматом и если чувствительность комплекса белков внутри клетки достаточно высока, то импульс возникнет. Рецепторы другого типа – NMDAR – работают далеко не так просто: они ведут себя по-разному в зависимости от того, есть в данный момент импульс в клетке или нет. Если нейрон находится в состоянии покоя, молекулы NMDAR ничего не могут сделать, даже если вокруг много глутамата. Однако в возбужденном нейроне (то есть в таком нейроне, который получает достаточно сигналов от всех своих синапсов для возникновения импульса) молекулы NMDAR способны реагировать на глутамат и направлять свои сигналы в клетку. Эти сигналы не активируют клетку напрямую, а локально изменяют систему AMPAR этого синапса, увеличивая силу сигнала, который может создать определенное количество глутамата. Именно рецепторы NMDAR лежат в основе гипотезы Хебба: они активны, если уже активны и принимающая клетка, и конкретный синапс (за счет наличия глутамата), и могут изменить силу синаптической связи, повысив чувствительность системы AMPAR (рис. 73).
Важность системы NMDAR для обучения была наглядно продемонстрирована в экспериментах с плодовыми мушками, запахами и электрическими разрядами. Были созданы генетически модифицированные плодовые мушки, у которых можно было в любой момент отключить производство белков NMDAR и затем снова включить его. Поскольку белки NMDAR довольно короткоживущие, их практически не остается в организме плодовой мушки уже через пятнадцать часов после отключения. У таких мушек способность ассоциировать запах с электрическими разрядами сильно нарушена. После возобновления производства NMDAR к мушкам возвращается способность «учиться» в обычном режиме.
Рис. 73. Влияние потенциал-чувствительного рецептора NMDAR на характер стимуляции принимающей клетки рецепторами AMPAR. Вентиль обозначает сети сигнальных белков и систем, контролирующих содержание AMPAR в мембране. Вместе они регулируют силу сигнала, который может подать система AMPAR при наличии конкретного количества глутамата
Хеббовская регулировка силы синаптических связей – один из возможных, но не единственный механизм обучения. Есть убедительные доказательства того, что и у человека, и у низших организмов сами нейронные структуры головного мозга могут меняться в зависимости от жизненного опыта. Один из самых изученных примеров относится к развитию зрительной системы. Давайте посмотрим, что происходит после того, как «карта» сетчатки глаза была в общих чертах нанесена на определенную область головного мозга (глава 13). Связь аксонов сетчатки и нейронов в мозге, возникшая под действием химических сигналов, довольно приблизительна. По мере создания синаптических связей в верхнем двухолмии и других областях мозга аксоны из определенной части глаза подключаются не только ко многим «правильным» клеткам (к которым подключаются также соседние аксоны), но и к нескольким «неправильным» клеткам. Отсутствие корректировки привело бы к затуманиванию зрения, и возможности такой удивительной системы, как человеческий глаз, остались бы нераскрытыми. Поэтому после рождения ребенка, когда глаза открываются, нейронная сеть корректируется за счет перестройки связей, связанной с активностью глаз. Этот механизм напоминает механизм Хебба, но на этот раз от совпадения импульса синапса и импульса принимающей клетки зависит не только сила синаптических связей, но и само их сохранение. Если, например, аксон из левой верхней части глаза образует синапс с нейроном, с которым уже соединены аксоны других клеток верхней левой части глаза, импульс будет возникать одновременно, потому что все они будут передавать информацию об одних и тех же событиях, попадающих в поле зрения. Совместное действие всех этих синапсов будет возбуждать принимающий нейрон, и все его синаптические соединения усилятся. По мере того как принимающая клетка будет передавать все больше импульсов, она будет все неохотнее сохранять слабые синапсы, которые никогда не «попадают в такт». Синапс, не синхронизированный с импульсами принимающей клетки, скорее всего, реагирует не на те стимулы, на которые реагируют большинство синапсов этой принимающей клетки. Это значит, что его аксон выходит из другой части глаза или его передающий нейрон чувствителен к какой-то другой части поля зрения. Как бы то ни было, в этой принимающей клетке такому синапсу не место. Отключая такие «рассинхронизированные» синапсы, принимающие клетки освобождаются от груза неправильных соединений. Свободный конец аксона может «попытать счастья» и подключиться к другой принимающей клетке (за счет химических сигналов, описанных в главе 13). Процесс повторяется до тех пор, пока аксон не найдет подходящий нейрон и не образует с ним синапс, синхронизированный с другими синапсами этого нейрона.
