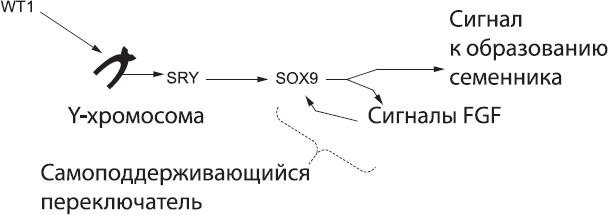
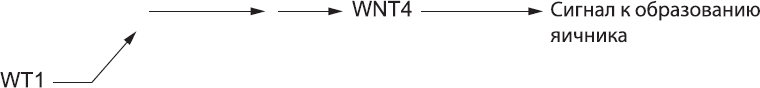Рис. 57. Биохимический каскад «самцовости», который активирует переключатель SOX9 – FGF
Важность петли SOX9 – FGF была подтверждена в ряде экспериментов над генетически модифицированными мышами. Эмбрионы мышей, в клетках гонад которых Sox9[209] полностью отсутствовал, формировали признаки, характерные для самок, несмотря на присутствие Sry, так как Sry не может влиять на организм без работоспособного Sox9.[210] Такой же эффект наблюдался после удаления сигнальной системы Fgf. Напротив, мыши, в гонадах которых экспериментально активировали экспрессию Sox9 независимо от присутствия Sry, развивались в самцов, даже если у них не было Y-хромосомы.[211]
Под действием продуктов генов, активированных белком SOX9, клетки, продуцирующие этот белок, начинают активно размножаться, а также приобретают характерную форму и биохимические свойства клеток семенников, которые обеспечивают образование сперматозоидов. После того как эти клетки окончательно выбрали путь развития, они заставляют перейти к формированию семенника и все остальные клетки гонады (семенник содержит большое количество трубочек, в толстых стенках которых в свое время будут формироваться из клеток зародышевой линии сперматозоиды).
Мы только что обсудили мужской путь развития. Чтобы проследить путь женского развития, следует вернуться к тому моменту, когда в гонаде, еще не определившейся с выбором пола, образовывался белок WT1. Он действует далеко не только на ген SRY, а вызывает также последовательную активацию ряда других генов, находящихся на обычных хромосомах, имеющихся у всех эмбрионов, независимо от того, имеется у них ген SRY или нет. Мы пока не знаем все детали этой последовательной активации, но известно, что через несколько часов клетки начинают продуцировать еще один сигнальный белок из известного нам семейства WNT (см. главы 7, 10 и 11). Это белок WNT4. При условии отсутствия сильных ингибиторов его функции этот белок заставляет клетки гонад формировать яичник (рис. 58).
Понятно, что у мужчин путь WNT4 должен как-то блокироваться, и петля FGF – SOX9, активируемая белком SRY в мужских половых железах, как раз и является мощным ингибитором функций WNT4. Поэтому общий принцип заключается в следующем: «по умолчанию» эмбрион следует по женскому типу развития под руководством WNT4, а если гены, характерные для мужского пути, успели активироваться, женский путь развития подавляется.[212]
Рис. 58. Женский путь развития: экспрессия WT1 в конечном итоге приводит к экспрессии WNT4, а он подает клеткам гонады сигналы к образованию яичника
Как мы помним, мужской путь развития снабжен «переключателем» – сигнальной петлей SOX9 – FGF, который гарантирует, что клетки не сойдут с выбранного пути из-за случайных колебаний в системе. Женский путь развития также снабжен специальным переключателем, благодаря которому слабая или случайная активация «мужских» генов не приводит к полному сумбуру. После того как сигнальный путь WNT4 активирован, он ингибирует «мужской» путь, гарантируя, что никакой всплеск активности «мужских» генов ни в какой клетке не переведет «переключатель» в «мужское» состояние. Поэтому выбор пола – это вопрос борьбы за ручку переключателя. Если SRY присутствует и вовремя включает петлю FGF – SOX9, то развитие автоматически идет по мужскому пути, а женский путь WNT4 находится в положении «выключено». Если же SRY отсутствует или по какой-то причине не активирует петлю FGF – SOX9 вовремя (у мышей речь идет примерно о шести часах), то запускается женский путь развития, а мужской путь переходит в положение «выключен» (рис. 59).
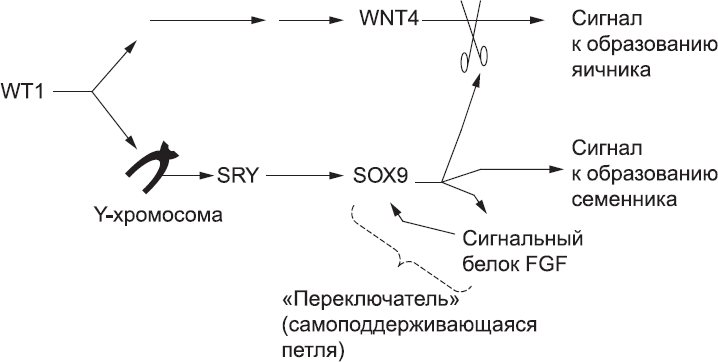
Рис. 59. Молекулярная логика определения пола. Появление в гонаде белка WT1 запускает последовательность событий, которая в конечном итоге приводит к синтезу WNT4, а он подает клеткам гонад сигнал к созданию яичника. Однако если у эмбриона есть Y-хромосома, WT1 также активирует находящийся на ней ген, который запускает производство белка SRY. Это стимулирует производство SOX9. Он имеет несколько функций: активирует сигнальный путь FGF, который поддерживает синтез SOX9, блокирует действие WNT4 («заглушая» сигнал к формированию яичников) и подает клеткам гонады сигнал формировать семенник
Важность сигнального пути WNT4 для выбора эмбрионом женского пути развития была показана во многих экспериментах над генетически модифицированными мышами. Мыши, в гонадах которых за счет какого бы то ни было генетического вмешательства был активирован сигнальный путь WNT4, развивались по женскому пути, даже если имели Y-хромосому.[213]
Если основанный на WNT4 «переключатель» в соматических клетках гонады находится в положении «женский путь развития», то они будут развиваться в клетки, обеспечивающие формирование яйцеклеток. Они посылают сигналы, благодаря которым остальные клетки половой железы образуют ткани яичника, а не семенника (яичники не состоят из многочисленных трубочек, а заполнены рыхлой тканью и группами клеток, которые собираются вокруг развивающихся яйцеклеток). Один из сигналов этих клеток подталкивает зародышевые клетки к особому типу деления – мейозу, – который имеет решающее значение для формирования как яйцеклеток, так и сперматозоидов. В организме мальчиков мейоз начинается только после наступления половой зрелости, но у девочек все клетки зародышевой линии вступают в мейоз практически в самом начале развития яичников: на момент рождения девочки все ее яйцеклетки находятся в состоянии мейоза. Хотя мейоз начинается еще до рождения, он, как ни странно, приостанавливается в период примерно с двенадцати до пятидесяти лет, и только несколько яйцеклеток возобновляют свое развитие каждый менструальный цикл. Такой характер развития имеет одно неприятное медицинское следствие. Развивающиеся яйцеклетки, сделавшие паузу в мейозе, крайне уязвимы к лекарственным препаратам, которые иногда используется в химиотерапии, и для больных раком девочек и молодых женщин ценой спасения может оказаться бесплодие. К счастью, был разработан метод, позволяющий удалять участки ткани яичников перед началом химиотерапии, замораживать их, а затем трансплантировать обратно, если женщина, достигнув детородного возраста, решит, что хочет завести детей.[214]
Мейоз, в отличие от обычного клеточного деления, не приводит к образованию двух дочерних клеток с теми же хромосомами, что были и у материнской. Обычная клетка человеческого тела перед делением имеет по две копии каждой хромосомы, одна из которых унаследована от матери, а другая – от отца. Есть две копии хромосомы 1, две копии хромосомы 2, и так далее. Если клетка женская, у нее есть две копии Х-хромосомы, а если клетка мужская, то у нее есть одна Х-хромосома и одна Y-хромосома (потому что, как я говорил выше, именно присутствие Y-хромосомы и белка SRY приводит к созданию мужского организма). При нормальном делении каждая хромосома копируется, и дочерние клетки получают тот же набор, что и материнская клетка, а вот при мейозе хромосомы распределяются между клетками таким образом, что одна из копий достается одной дочерней клетке, а вторая – другой. Поэтому в конечном итоге дочерние клетки обладают только одной копией каждой хромосомы. Это именно то, что нужно яйцеклетке или сперматозоиду – когда в процессе оплодотворения они сольются, у оплодотворенной яйцеклетки будет нормальное количество хромосом. Все яйцеклетки, образованные в процессе мейоза в организме женщины, несут исключительно Х-хромосомы. Однако в мужском организме половина сперматозоидов несет Х-хромосому, а вторая половина – Y-хромосому. Если яйцеклетку оплодотворит сперматозоид с X-хромосомой, у эмбриона будут две Х-хромосомы и ни одной Y-хромосомы, и родится девочка. Если же яйцеклетку оплодотворит сперматозоид с Y-хромосомой, то эмбрион получит Y-хромосому (а с ней и ген SRY), и родится мальчик. Примерное соотношение мужчин и женщин в человеческой популяции – 50/50[215] – непосредственно связано с характером распределения хромосом при мейозе.