Начну с ряда бесспорных фактов. Во-первых, конечности удлиняются постепенно, и основная пролиферация клеток происходит в так называемой зоне прогресса, расположенной в дистальной части конечности. По мере удлинения конечности зона прогресса оставляет позади себя клетки (так оставляет за собой след ползущая улитка), которые дифференцируются и формируют структуры конечностей. Во-вторых, клетки на обоих концах развивающейся конечности подвергаются влиянию разных наборов сигнальных молекул (или, во всяком случае, разного соотношения сигнальных молекул). Клетки эктодермы на самом конце зачатка конечности выделяют белки FGF.[188],[189] На плечевом конце в зачаток конечности поступает из туловища другая молекула – ретиноевая кислота (о ней уже шла речь, когда мы обсуждали образование сомитов). В-третьих, плечевые кости закладываются раньше, чем кости предплечья, а те, в свою очередь, – раньше, чем кости кисти.
Тот факт, что клетки, проводящие разное время в зоне прогресса, в конечном итоге создают разные части руки, как раз и лежит в основе одной из самых спорных современных моделей определения положения элементов конечностей.[190] Согласно этой модели, время, проведенное в зоне прогресса, само по себе является сигналом, и чем больше времени клетки провели в ней, тем больше признаков, характерных для пальцев, приобретет сформированная ими структура (рис. 52). Клетки, которые рано покинули зону прогресса, образуют плечо, а клетки, покинувшие ее немного позже, – локоть. Клетки, запоздавшие еще сильнее, образуют предплечье, запястье и кисть, а клетки, которые, по сути, так и не покинули зону прогресса, – кончики пальцев. Эта изящная модель предсказывает, что если вынудить клетки провести в зоне прогресса аномально долгое время, то плечевой ткани в конечности будет мало, а ткани кисти – слишком много. Пытаясь создать именно такие условия, исследователи облучали зачаток конечности цыпленка рентгеновскими лучами. Под действием облучения многие клетки в зоне прогресса погибли, а те, которые выжили, провели в этой зоне больше времени, чем обычно, восстанавливая свою численность за счет пролиферации; лишь после этого некоторые из них смогли покинуть ее. В результате действительно образовалась конечность без плечевой кости, но со структурами кисти, в полном соответствии с предсказанием временной модели.[191]
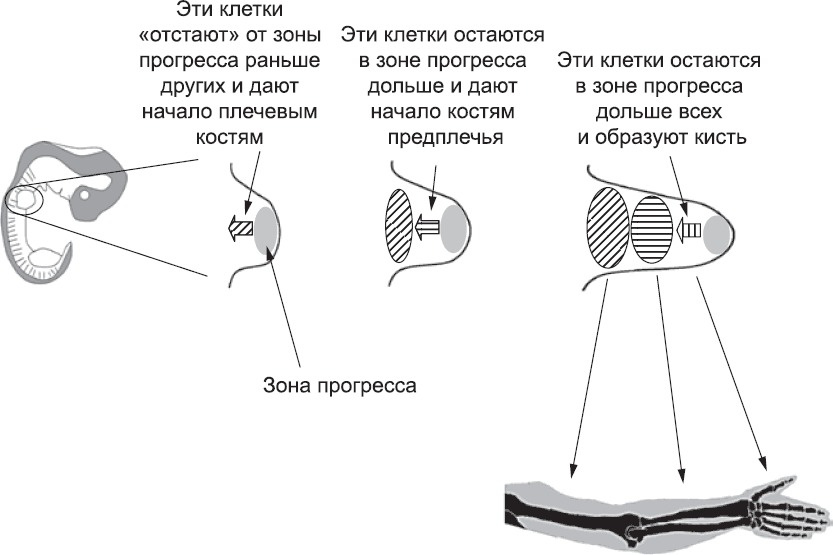
Рис. 52. Временная модель развития конечности: по мере удлинения зачатка конечности активно размножающиеся клетки «отстают» от зоны прогресса в разное время. Согласно этой модели, кончики пальцев руки, например, будут образованы клетками, которые провели больше всего времени в зоне прогресса. На схеме изображены только три стадии, соответствующие трем крупным отделам руки, но эта модель работает и для более детального уровня дифференцировки структур конечности
Проблема описанного выше эксперимента в том, что отложенный выход выживших клеток из зоны прогресса был лишь косвенным эффектом воздействия рентгеновских лучей. Их непосредственный эффект выражался в гибели многих других клеток. Резонно предположить, что отсутствие плечевых костей с тем же успехом могло объясняться разной чувствительностью разных типов клеток к рентгеновскому излучению. Недавно этот эксперимент был повторен[192] с использованием современных методов, позволяющих отследить экспрессию генов, и выяснилось, что рентгеновские лучи никак не влияют на клетки, находящиеся на ранних этапах специализации в клетки плечевых структур. Изучение убивало только клетки, находившиеся в процессе создания видимых структур плеча. Таким образом, эксперимент, задумывавшийся для подтверждения временной модели, не оправдал надежд. Нельзя сказать, что эта модель была опровергнута, просто приходится признать, что она не столь хорошо подкреплена доказательствами, как считалось раньше.
Другая модель определения положения структур конечности никак не связана со временем. Она основана на существовании двух противоположных градиентов концентрации – белков FGF от кончиков пальцев к плечу и ретиноевой кислоты в обратном направлении. Суть в следующем: при таком распространении сигнальных молекул клетки в разных точках конечности подвергаются воздействию разного соотношения концентраций ретиноевой кислоты и FGF, и эта разница определяет различия в их дальнейшем поведении (рис. 53). Поскольку сигнальные молекулы могут распространяться лишь на небольшие расстояния, большинство версий этой модели постулируют, что клетки приобретают отличительные черты на ранних стадиях, когда зачаток конечности небольшой, даже если их дифференциация проявляется гораздо позже. Основная идея этой модели подтверждается экспериментами, в которых клетки извлекали из ранних зачатков конечностей и выращивали в чашках Петри. Если добавить в такую культуру чистые сигнальные белки, все клетки получат их в одинаковой концентрации.[193] Под действием сигнальных белков FGF, которые в норме поступают из дистальной части зачатка конечности, в клетках подавлялась экспрессия генов, связанных с развитием плеча, и увеличивалась сначала экспрессия генов, связанных с развитием предплечья, а затем генов, связанных с развитием кисти. Ретиноевая кислота, источником которой в норме является плечевая часть руки, стимулировала формирование плечевых структур, даже если одновременно с ретиноевой кислотой в культуру добавляли белки FGF. Похожие результаты были получены и в серии экспериментов, в которых клетки зачатка конечности трансплантировали в другие участки эмбриона, богатые FGF или ретиноевой кислотой.[194] В частности, эти эксперименты ясно показали, что для формирования верхней части руки нужна ретиноевая кислота или эквивалентная ей сигнальная молекула, поступающая со стороны туловища.[195]
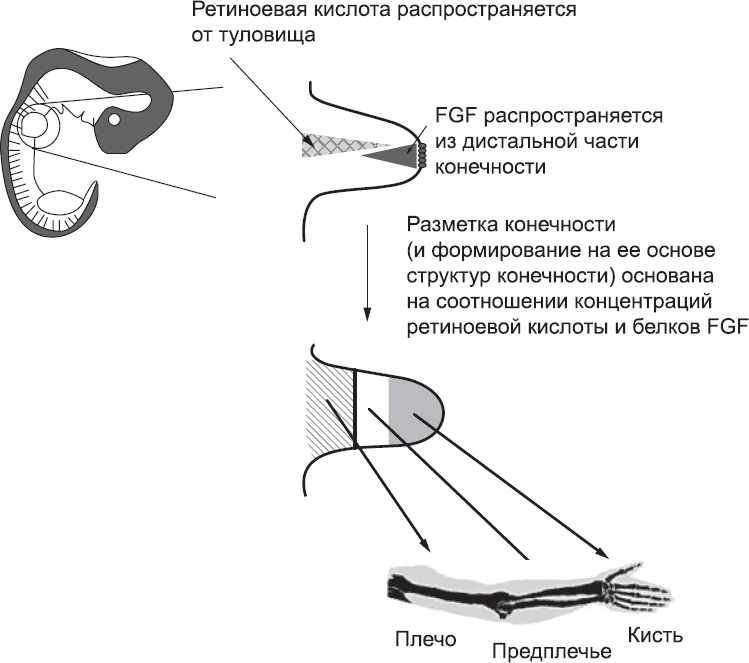
Рис. 53. Модель, основанная на соотношении концентраций сигнальных молекул. Противоположные градиенты концентрации – белка FGF из дистальной части конечности и ретиноевой кислоты, поступающей из туловища, – активируют разные паттерны экспрессии генов. Клетки, которые подвергаются воздействию высокой концентрации FGF и низкой концентрации ретиноевой кислоты, образуют структуры кисти, а клетки, на которые белок FGF действует в низкой концентрации, а ретиноевая кислота – в высокой, образуют структуры плеча. Точные уровни концентрации FGF и ретиноевой кислоты определяют, образует ли клетка структуры, характерные для кончиков пальцев или для основной части кисти, для локтя или для плеча
Если модель, основанная на соотношении концентраций сигнальных молекул, верна, то добавление дополнительной порции FGF в зачаток конечности должно вызывать образование более обширной зоны кисти или пальцев в ущерб остальной части руки. Однако этого не наблюдается – зона клеток, идущая на формирование кисти, имеет такой же размер, что и обычно.[196] Это говорит о том, что модель, основанная на соотношении концентраций сигнальных молекул, не вполне верна, во всяком случае для развития области кисти.
Итак, на данный момент у нас есть по меньшей мере две основные модели развития конечности. Каждая из них подтверждается рядом фактов, но расходится с результатами как минимум одного эксперимента. Когда в науке возникает проблема такого рода, полезно подумать, нет ли каких-то неочевидных допущений, присущих обеим моделям. В нашем случае такое скрытое допущение есть, и оно заключается в том, что разметка всей конечности от плеча до кончиков пальцев осуществляется с помощью одного и того же механизма. Возможно, это неверно. В эволюции сложная конечность возникла не сразу. В грудных плавниках лопастеперых рыб, например, есть структуры, гомологичные плечу и предплечью, но нет ничего, что соответствовало бы кисти. Значит, развитие кисти «добавилось» к развитию конечности позже. Это подтверждается тем, что при развитии конечностей наземных животных экспрессия генов проходит те же основные стадии, что и у рыб, а затем появляется совершенно новый, отсутствующий у рыб этап. Более примитивные бесчелюстные рыбы имеют (или, в случае вымерших видов, имели) даже более примитивные плавники.[197] Таким образом, возможно, что части конечности, расположенные на разных уровнях оси «плечо – кончики пальцев», размечаются за счет разных механизмов (рис. 54). Тогда возможно, что модель, основанная на соотношении сигнальных молекул, правильно объясняет, скажем, особенности формирования плечевой части – сигнальная молекула (ретиноевая кислота), источником которой является туловище, «защищает» прилегающую часть руки от превращения в какую-либо другую область конечности. А особенности образования предплечья и кисти, которые после удлинения зачатка конечности оказываются за пределами распространения сигналов от туловища, возможно, верно объясняются временной моделью. Тогда становится понятно, почему повышенная доза белка FGF не увеличивает размер кисти в ущерб плечу.