Такое чередование рутинных событий со скачкообразными изменениями свойственно не только жизни взрослого человека, но и жизни эмбриона. На ранних этапах развития эмбриона, описанных в предыдущих двух главах, происходило всего лишь простое деление клеток. Затем – на первый взгляд, внезапно – начались новые процессы, связанные с формированием клеточных слоев, заметно отличающихся друг от друга. Получившаяся в результате структура – по сути, наполненная жидкостью сфера, которая разделена на две камеры парой дисков (рис. 9), – по-прежнему далека от того, чтобы в ней можно было опознать что-то человекоподобное. Скажите кому-нибудь, что из этой пары дисков получится животное, и он, вероятно, предположит, что это будет медуза. По крайней мере, ее колокол, как и диски эмбриона, радиально симметричен, и у нее есть верх и низ, но нет оси, разделяющей тело на половинки (см. рис. 9). Тем не менее эмбрион уже успел подготовиться к масштабной реорганизации, после которой он уже будет напоминать человека. Этот процесс, занимающий всего пару дней, называется гаструляция.
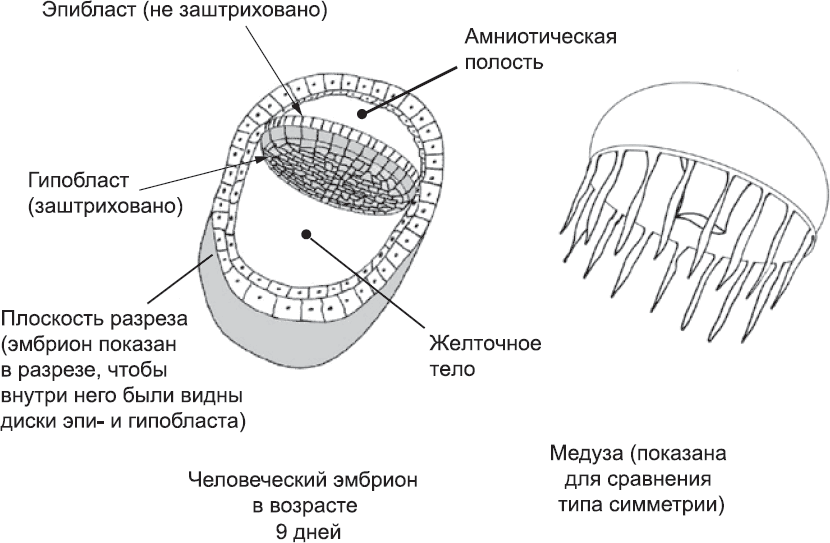
Рис. 9. Простая радиальная симметрия зародышевого диска человеческого эмбриона в сравнении с аналогичной симметрией медузы
Чтобы подойти к теме гаструляции, можно было бы сначала рассмотреть строение примитивного организма, который образуется в результате данного процесса, а затем посмотреть, какой вклад в его образование вносит каждый этап гаструляции. Это, пожалуй, был бы самый рациональный и простой подход к описанию гаструляции, однако он может создать ложное впечатление о том, что клетки имеют какое-то представление об анатомии организма, в строительстве которого участвуют. На самом же деле развитие организма основано вовсе не на том, что клеткам известны законы развития (понять эти законы трудно даже нам, с нашим мозгом из миллионов клеток), а на том, что клетки автоматически откликаются на изменения окружающей их среды. Поэтому в этой главе мы, следуя логике развития, сначала рассмотрим поведение клеток развивающегося организма и только потом посмотрим, к чему оно приводит.
Прежде чем начать рассказ, хочу сделать важное предупреждение: изучать процесс гаструляции у человека крайне сложно, поэтому почти все, что описано в этой главе, основано на исследованиях животных. Существуют строгие правовые ограничения относительно выращивания человеческих эмбрионов в лабораторных условиях, и изучать эмбрионы на стадии гаструляции нельзя (почему – я расскажу чуть позже). Основная последовательность анатомических изменений человеческого эмбриона известна. Эти данные были получены в исследованиях немногочисленных (и очень ценных) эмбрионов, извлеченных при посмертном вскрытии или при удалении матки у женщин, которые чаще всего даже не знали о том, что беременны. Гаструляция начинается примерно через пятнадцать дней после оплодотворения, то есть примерно тогда, когда женщина ожидает начала менструации. Некоторым из этих эмбрионов более ста лет, но они до сих пор хранятся в музеях под особым присмотром, потому что новый материал такого рода поступает крайне редко. Исследования проводятся главным образом на мышах и курицах, а процесс гаструляции у этих животных значительно отличается от гаструляции у человека. Цыпленок развивается в яйце, а не в матке; эпибласт и гипобласт мыши имеют форму чаши, а не диска. Эти особенности могут играть существенную роль при развитии. Поэтому реконструировать механизмы гаструляции человека на основе данных, полученных при изучении модельных животных, рискованно – слишком мало у нас информации, и мы можем неверно интерпретировать какие-либо детали процесса.
Отправной точкой для гаструляции является эмбрион в том состоянии, в котором мы оставили его в конце главы 3. К этому времени уже сформировался ряд вспомогательных тканей, таких как плацента, а также два заполненных жидкостью внутренних пространства: амниотическая полость и желточный мешок. Между этими полостями лежат два диска, расположенные один над другим: гипобласт и эпибласт. Из гипобласта образуются дополнительные вспомогательные ткани, а из эпибласта – сам плод. Ни один из этих двух дисков не имеет каких-либо явных особенностей, которые позволяли бы отличить один его край от другого (см. рис. 9).
Первое изменение (судя по данным, полученным на животных) происходит в гипобласте. У клеток, расположенных в середине этого диска,[36] включаются новые гены, в том числе и ген ДНК-связывающего белка, который называется HEX.[37] Что является сигналом к этому изменению, пока неясно. Возможно, что все клетки гипобласта изначально готовы к нему, но большинство из них ингибируется сигнальным белком, который синтезируется тканями, окружающими гипобласт.[38] Достаточно далеко от источника этого ингибитора находятся только клетки в центре диска, поэтому они могут избежать его влияния и запустить экспрессию гена HEX. Этот механизм – только предположение, но активация гена HEX – факт. Клетки, в которых экспрессируется HEX, уходят из центра диска – они перемещаются, расталкивая соседние клетки, и собираются в одной точке на краю диска гипобласта[39],[40],[41] (рис. 10). Все еще непонятно, даже в случае мышей, что же особенного в этой точке диска, что клетки мигрируют именно туда. У «низших» животных расположение похожей «особой» точки обусловлено начальными условиями развития. В некоторых случаях положение этой точки определяется переходящей к зародышу асимметрией распределения молекул, заложенных в яйцеклетку материнским организмом. В других случаях в этой точке располагаются полярные тельца, «побочные продукты» клеточных делений, предшествующих формированию яйцеклетки. У некоторых организмов эту точку, по-видимому, маркирует место проникновения сперматозоида. Весьма вероятно, что у млекопитающих это тоже так, и есть указания на то, что эмбрионы мышей асимметричны уже на самых ранних стадиях развития.[42] Соответствующих данных по эмбрионам людей, разумеется, нет. Это очень досадно, потому что скопление в одном месте клеток, у которых экспрессируется HEX, приводит к исключительно важному результату – одно конкретное место на краю диска гипобласта становится не таким, как другие. Иными словами, это первый шаг в сторону от простой радиальной симметрии эмбриона (рис. 10).
Достигнув края диска гипобласта, группа клеток, экспрессирующих ген HEX, становится новой структурой ПВЭ.[43] Клетки ПВЭ начинают секретировать собственные сигнальные белки. Эти белки, способные распространяться на небольшие расстояния, вполне могут достигнуть эпибласта, лежащего непосредственно над гипобластом, явно в пределах дистанции распространения сигнала.[44] На этом этапе клетки эпибласта уже реагируют на сигналы, поступающие от вспомогательных тканей эмбриона. Эти сигналы подготавливают их к тому, чтобы создать структуры, характерные для задней части организма. Если бы больше ничего не происходило, весь эпибласт формировал бы структуры задней части, и это имело бы катастрофические последствия для эмбриона. Сигналы, поступающие от ПВЭ, уравновешивают эту тенденцию, запуская процессы создания тканей головы.[45] Если ПВЭ не будет синтезировать сигнальные белки, эмбрион не сможет правильно сформировать голову. Таким образом, положение ПВЭ в уникальной области на краю диска гипобласта диктует полярность вышележащего эпибласта: сторона, ближайшая к ПВЭ, сформирует голову, а удаленная от нее – заднюю часть тела эмбриона. Только небольшая часть эпибласта удалена от ПВЭ настолько, что вообще не получает от нее сигналов. По-видимому, это очень важно для локализации области, в которой происходят первые заметные изменения эпибласта. В этом месте клетки эпибласта переключаются на образование сигнального белка, который привлекает окружающие клетки – они мигрируют по направлению к источнику этого белка.[46] В результате такого перемещения клеток образуется важная структура со скромным названием – первичная полоска (см. рис. 10).