303. Tоnini G. — Boll. soc. ital. Biol, sperim., 1955, v. 31, p. 766.
304. Tоnini G. — Boll. soc. ital. Biol, sperim., 1958, v. 34, p. 793.
305. Truitt E. — J. Nerv. Ment. Dis., 1958, v. 126, p. 184.
306. Truitt E., Ebersberge E. — Science, 1962, v. 135, p. 105.
307. Twarog M., Page I. — Amer. J. Physiol., 1953, v. 175, p. 157.
308. Udenfriend S., Bogdanski D., Weissbach H. — Fed. Proc., 1956, v. 15, p. 493.
309. Udenfriend S., Titus E., Weissbach H. — J. biol. Chem., 1955, v. 216, p. 499.
310. Udenfriend S., Titus E., Weissbach H., Peterson R. — J. Biol. Chem., 1956, v. 219, p. 335.
310. Udenfriend S., Weissbach H., Bogdanski D. — Ann. N. Y Acad. Sci., 1957, v. 66, p. 602.
312. Udenfriend S., Weissbach H., Bogdanski D. — J. Biol. Chem., 1957, v. 224, p. 803.
313. Udenfriend S., Weissbach H., Sjordsma A. — Science, 1956, v. 123, p. 669.
314. Valcourt A. — Arch. Neurol. Psychiat., 1959, v. 81, p. 292.
315. Valdecasas F., Calvet F., Cuenca E., Salvá J. — Arzneimittel-Forsch., 1956, Bd. S. 594.
316. Vassilia G., Costa E., coll. — Amer. J. Psychiat., 1961, v. 117, p. 1121.
317. Ventra D., Serra C. — Acta Neurol., 1956, p. 1112, p. 935.
318. Vincet D., Segonzac G. — Thérapie, 1960, v. 15, p. 914.
319. Voelkel A. — Confin. Neurol., 1958, v. 18, p. 144.
320. Voelkel A. — Psychiat. Neurol., 1960, Bd. 140, S. 36.
321. Wada J. — Science, 1961, v. 134, p. 1688.
322. Walaszek E., Abood L. — Proc. Soc. exper. Biol. Med., 1959, v. 101, p. 37.
323. Warner R., Kirschner P., Warner G. — J. A. M. A., 1961, v. 178, p. 1175.
324. Weissbach H., Bogdanski D., coll. — J. Biol. Chem., 1957, v. 227, p. 617.
325. Weissbach H., Bogdanski D., Udenfriend S. — Arch. Biochem. Biophysics, 1958, v. 73, p. 492.
326. Weissbach H., Lovenberg W., coll. — J. Pharmacol. exper. Therap., 1961, v. 131, p. 26.
327. Winters W. — Marquette Med. Rev., 1959, v. 24, p. 204.
328. Woolley D. — Rend. Ist. super. sanita, 1956, v. 19. Suppl. p. 231
329. Woolley D. — Science, 1957, v. 125, p. 752.
330. Woolley D. — Res. Publ. Ass. Nerv. Ment. Dis., 1958, v. 36, p. 381,
331. Woolley D. — В кн. Pennes H., Psychopharmacology, London, 1958, p.152.
332. Woolley D. — Dis. Nerv. Syst., 1960, v. 21, Section II, p. 87.
333. Woolley D., Shaw E. — Science, 1954, v. 119, p. 587.
334. Woolley D., Shaw E. — Brit. Med. J., 1954, v. 2, p. 122.
335. Woolley D., Shaw E. — Ann. N. Y. Acad. Sci., 1957, v. 66, p. 649.
336. Yen C., Stanger R., Millman N. — Arch. internet, pharmacodyn., 1959, v. 123, p. 179.
337. Zaimis E. — Progr. med., 1958, v. 86, p. 411.
338. Zanowiak P., Rodman M. — J. Amer. Pharmacol. Assoc., Scient. Ed., 1959, v. 48, p. 165.
339. Zeller E. — J. clin. exper. Psychopathol., 1958, v. 19, Suppl. I, p. 106.
340. Zeller E., Bernsohn J., Inskip W., Lauer J. — Naturwissenschaften, 1957, v. 44, p. 427.
341. Zettler G., Sсhlоsser L. — Arch. exper. Path. u. Pharmak., 1954, Bd. 222, S. 345.
342. Zuanazzi G., Rognoni F. — Atti. Soc. Lombarda sci. med. biol., 1959, v. 14, p. 305.
Глава 2
ОБМЕН АДРЕНАЛИНА И ПСИХИЧЕСКИЕ НАРУШЕНИЯ
Адреналин и его роль как медиатора симпатической нервной системы известны уже с начала нынешнего столетия. Вопросу о синтезе и расщеплении адреналина посвящено огромное количество работ. Изложение этих данных приведено в соответствующих руководствах и не является нашей задачей. Мы лишь кратко приведем те основные выводы, которые имеют значение для оценки гипотез, касающихся роли адреналина в возникновении психических заболеваний.
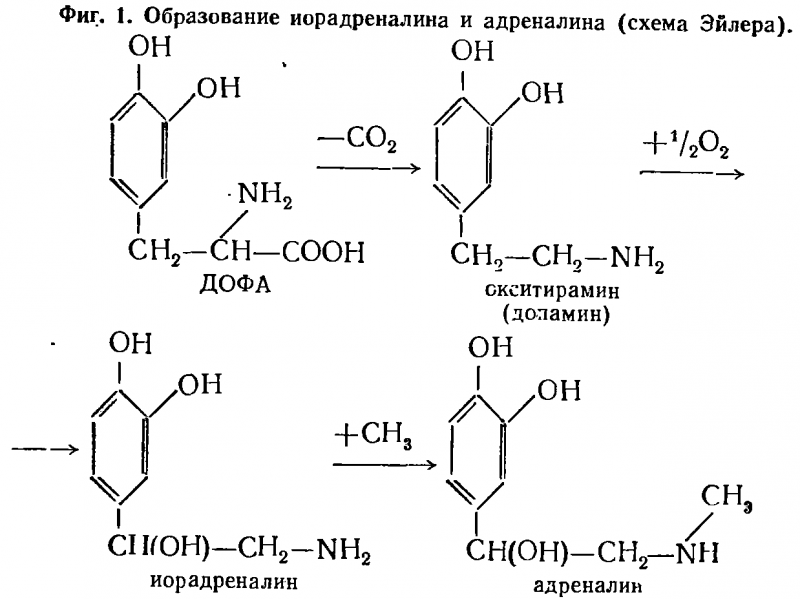
Адреналин и норадреналин синтезируются из более простых ароматических аминов и аминокислот, вероятно, из фенилаланина. Непосредственными предшественниками адреналина являются диоксифенилаланин (ДОФА) и образующийся из него окситирамин (более известный под названием допамин) который, окисляясь, переходит в норадреналин и посредством метилирования — в адреналин (фиг. 1).
В головном мозгу норадреналин обнаружен в 1946 году фон Эйлером и в 1950 году Гольцем. В экспериментах на животных было показано, что норадреналин в различных отделах головного мозга распределяется неравномерно. Наиболее высокая его концентрация обнаружена в гипоталамической области. Относительно высокие концентрации норадреналина обнаружены также в сером веществе вокруг Сильвиева водопровода, в ряде других структур среднего мозга, обонятельном мозге, несколько меньшие — в ретикулярной формации ствола головного мозга, на дне 4-го желудочка в медиальных ядрах таламуса. В остальных структурах головного мозга норадреналин обнаружен лишь в виде следов. Допамин обнаружен в покрышке и в хвостатом ядре, где он, по-видимому, играет физиологическую роль, в остальных участках мозга он, очевидно, быстро превращается в норадреналин и концентрация его незначительна. С возрастом у животных возрастает как абсолютная, так и относительная (в расчете на 1 г вещества) концентрация норадреналина в головном мозгу. Норадреналин считают основным медиатором симпатической нервной системы в головном мозгу, где его концентрация по разным данным в 3—10 раз превосходит концентрацию адреналина. Однако имеются указания, что адреналин действует на нервную систему в несколько раз сильнее, чем норадреналин (при равных дозах). В мозгу не найдены ферменты, участвующие в образовании ДОФА, как и 5-гидрокситриптофана (5-ГТФ), но обнаружены декарбоксилазы, превращающие ДОФА в норадреналин (и 5-ГТФ в серотонин), как упоминалось в разделе, посвященном серотонину, ДОФА и ГТФ-декарбоксилазы идентичны. Очевидно, ДОФА и 5-ГТФ образуются вне мозга, а затем, проникая через гемато-энцефалический барьер, превращаются в серотонин и норадреналин, которые фиксируются тканями и фармакологически неактивны. Существование связанной и свободной формы не доказано, но весьма вероятно.
Согласно схеме, предложенной Коста и сотр., оба основных моноамина — серотонин и норадреналин — содержатся в нервной клетке в виде скоплений, резервного и мобильного, между которыми поддерживается постоянное равновесие. Эти скопления отделены липоидной мембраной от части клетки, в которой находится инактиватор моноаминов — МАО, а также рецепторы, воздействуя на которые они осуществляют свою основную функцию — повышение симпатического (норадреналин) или парасимпатического (серотонин) тонуса. Под влиянием нервного импульса моноамины проникают через мембрану, воздействуют на рецепторы и разрушаются МАО, проникновение через мембрану осуществляется также за счет процесса диффузии, подчиняющейся физико-химическим законам. Согласно этой схеме активную роль играют только свободные моноамины. Различные гипотезы, посвященные роли норадреналина и адреналина при психических заболеваниях учитывают только их роль как медиаторов симпатической нервной системы, хотя катехоламины оказывают влияние на ряд обменных процессов, происходящих в организме, в частности на углеводный обмен нервной клетки.
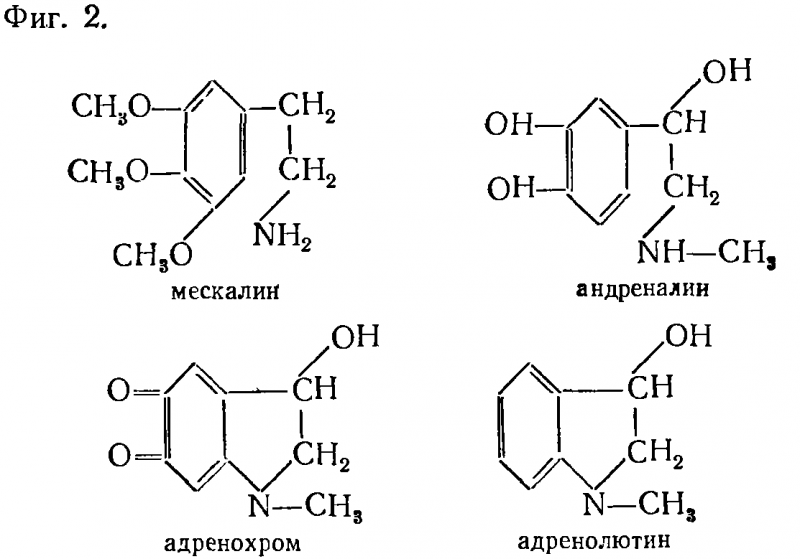
Адреналин по своему строению напоминает индол с незамкнутым кольцом и по своему строению близок к психотомиметическому средству — мескалину. На фиг. 2 приведены формулы адреналина и мескалина и двух метаболитов адреналина — адренохрома и адренолютина. Этим двум метаболитам придается наибольшее значение в генезе психических нарушений.
В организме адреналин быстро расщепляется. В отечественных руководствах выделено 3 основных пути обмена адреналина:
1) Окислительное дезаминирование под влиянием моноаминоксидазы с образованием 3, 4-диоксиванилиновой кислоты.
2) Образование ортохинона, превращающегося в адренохром и лейкоадренохром и, в конечном итоге, — в меланин.
3) Образование эфиров серной кислоты.