Из 64 кодонов у бактерий и фагов 3 кодона — УАА, УАГ и УГА — не кодируют аминокислот; они служат сигналом к освобождению полипептидной цепи с рибосомы, т. е. сигнализируют о завершении синтеза полипептида. Их назыают терминирующими кодонами. Существуют также 3 сигнала о начале синтеза — это т. н. инициирующие кодоны — АУГ, ГУГ и УУГ, — которые, будучи включенными в начале соответствующей информационной РНК (и-РНК), определяют включение формилметионина в первое положение синтезируемой полипептидной цепи. Приведённые данные справедливы для бактериальных систем; для высших организмов многое ещё не ясно. Так, кодон УГА у высших организмов может быть значащим; не совсем понятен также механизм инициации полипептида.
Реализация Г. к. в клетке происходит в два этапа. Первый из них протекает в ядре; он носит название транскрипции и заключается в синтезе молекул и-РНК на соответствующих участках ДНК. При этом последовательность нуклеотидов ДНК «переписывается» в нуклеотидную последовательность РНК. Второй этап — трансляция— протекает в цитоплазме, на рибосомах; при этом последовательность нуклеотидов и-РНК переводится в последовательность аминокислот в белке: этот этап протекает при участии транспортной РНК (т-РНК) и соответствующих ферментов (см. Белки, раздел Биосинтез).
Полный «словарь» генетического кода для аминокислот
| Первая «буква» | Вторая буква | Третья «буква» |
| У | Ц | А | Г |
| У | УУУ | Фенила- ланин | УЦУ | Серин | УАУ | Тирозин | УГУ | Цистин | У |
| | УУЦ | УЦЦ | УАЦ | УГЦ | Ц |
| | УУА | Лейцин | УЦА | УАА | Конец синтеза | УГА | Конец синтеза | А |
| | УУГ* | УЦГ | УАГ | УГГ | Триптофан | Г |
| Ц | ЦУУ | Лейцин | ЦЦУ | Пролин | ЦАУ | Гистидин | ЦГУ | Аргинин | У |
| | ЦУЦ | ЦЦЦ | ЦАЦ | ЦГЦ | Ц |
| | ЦУА | ЦЦА | ЦАА | Глутамин | ЦГА | А |
| | ЦУГ | ЦЦГ | ЦАГ | ЦГГ | Г |
| А | АУУ | Изолей- цин | АЦУ | Треонин | ААУ | Аспарагин | АГУ | Серин | У |
| | АУЦ | АЦЦ | ААЦ | АГЦ | Ц |
| | АУА | АЦА | ААА | Лизин | АГА | Аргинин | А |
| | АУГ* | Метионин | АЦГ | ААГ | АГГ | Г |
| Г | ГУУ | Валин | ГЦУ | Аланин | ГАУ | Аспараги- новая кислота | ГГУ | Глицин | У |
| | ГУЦ | ГЦЦ | ГАЦ | ГГЦ | Ц |
| | ГУА | ГЦА | ГАА | Глутами- новая кислота | ГГА | А |
| | ГУГ* | ГЦГ | ГАГ | ГГГ | Г |
* В начале цепи и-РНК данный кодон определяет начало синтеза полипептидной цепи и кодирует аминокислоту формилметионин. От готовых полиплоидных цепей формильная группа или вся аминокислота может быть отщеплена с помощью соответствующих ферментов.
Лит.: Общая природа генетического кода для белков, в сборнике: Молекулярная генетика, пер. с англ., М., 1963; Крик Ф., Генетический код (I), в кн.: Структура и функция клетки, пер. с англ., М., 1964, с. 9—23; Ниренберг М., Генетический код (II), там же, с. 24—41; Хэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Хартман Ф., Саскайнд З., Действие гена, пер. с англ., М., 1966; Бреслер С. Е., Введение в молекулярную биологию, 2 изд., М. — Л., 1966; Ингрэм В., Биосинтез макромолекул, пер. с англ., М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967; Сойфер В. Н., Молекулярные механизмы мутагенеза, М., 1969; Дубинин Н. П.. Общая генетика, М., 1970.
Н. П. Дубинин, В. Н. Сойфер.
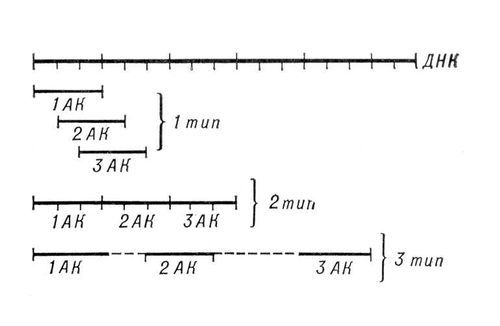
Модели генетического кода: 1-й тип — перекрывающийся код без запятых; 2-й тип — неперекрывающийся код без запятых; 3-й тип — код с «промежутками», т. е. код с запятыми.
Генетический полиморфизм
Генети'ческий полиморфи'зм, сосуществование в пределах популяциидвух или нескольких различных наследственных форм, находящихся в динамическом равновесии в течение нескольких и даже многих поколений. Чаще всего Г. п. обусловливается либо варьирующими давлениями и векторами (направленностью) отбора в различных условиях (например, в разные сезоны), либо повышенной относительной жизнеспособностью гетерозигот. Один из видов Г. п. — сбалансированный Г. п. — характеризуется постоянным оптимальным соотношением полиморфных форм, отклонение от которого оказывается неблагоприятным для вида, и автоматически регулируется (устанавливается оптимальное соотношение форм). В состоянии сбалансированного Г. п. у человека и животных находится большинство генов. Различают несколько форм Г. п., анализ которых позволяет определять действие отбора в природных популяциях.
Лит.: Тимофеев-Ресовский Н. В., Свирежев Ю. М., О генетическом полиморфизме в популяциях, «Генетика», 1967, № 10.
Генетическое действие излучений
Генети'ческое де'йствие излуче'ний, радиационный мутагенез, возникновение наследственных изменений (мутаций) при облучении организмов. Г. д. и. — важная часть биологического действия ионизирующих излучений, исследуемая радиационной генетикой. Первые стабильные «радиорасы» у дрожжей получены советскими биологами Г. А. Надсоном и Г. С. Филипповым (1925); данные о повышении частоты мутаций у дрозофилы при рентгеновском облучении опубликованы американским генетиком Г. Мёллером (1927). Мутагенный эффект вызывают все типы ионизирующих излучений, а также ультрафиолетовые лучи, если их действию подвергаются наследственные структуры любых организмов — от вирусов и бактерий до высокоорганизованных животных, включая человека; при этом у сложных организмов мутации могут возникать как в половых клетках — гаметах, так и в клетках тела — соматических. Облучение может вызывать все типы мутаций (генные, хромосомные, геномные и цитоплазматические). В определенном интервале доз частота мутаций возрастает пропорционально дозе облучения; при увеличении дозы выше некоторого значения линейность кривых, описывающих зависимость частоты мутаций от дозы, нарушается. Вновь возникающие мутации являются обычно рецессивными (см. Рецессивность) и вредными. Повышение радиоактивного фона ведёт к накоплению в популяциях организмов, в том числе и человека, скрытых вредных мутаций.
Важное практическое применение Г. д. и. — радиационная селекция, т. е. отбор хозяйственно-ценных мутаций, получаемых главным образом у культурных растений и промышленных микроорганизмов в результате их облучения. Выведенные таким способом новые сорта овса, ячменя, гороха, арахиса, плодовых и декоративных культур и др. уже занимают большие посевные площади. Многие высокопродуктивные промышленные штаммы микроорганизмов — продуцентов антибиотиков, витаминов, аминокислот — также получены путём радиационного мутагенеза.
Лит.: Итоги науки. Биологические науки, т. 3 — Ионизирующие излучения и наследственность, М., 1960; Астауров Б. Л., Функциональный принцип в оценке относительной значимости радиационных поражений ядра и цитоплазмы, в сборнике: Первичные механизмы биологического действия ионизирующих излучений, М., 1963; Ли Д. Э., Действие радиации на живые клетки, пер. с англ., М., 1963; Алиханян С. И., Селекция промышленных микроорганизмов, М., 1968; Тимофеев-Ресовский Н. В., Иванов В. И., Корогодин В. И., Применение принципа попаданий в радиобиологии, М., 1968.