В полногеномном взаимном обмене участвуют два целых генома. После обмена гомологичными участками остается по-прежнему два целых генома. Это совсем другое дело! Теперь клетка, в которой находятся эти геномы, может просто поделиться, отправив в каждую из дочерних клеток по одному геному[45]. Еще и на репликации сэкономит, а главное – никто не обижен, никто не идет на выброс. Следовательно, нет и оснований для распространения мутаций, защищающих гены от рекомбинации.
Только переход к полногеномному взаимному генетическому обмену мог сделать интенсивную межорганизменную рекомбинацию эволюционно стабильной, т. е. защищенной от эгоистичных аллелей, подобных tr−. Именно это, как мы думаем, и произошло у древних эукариот.
Промежуточным звеном на пути от ГПГ к амфимиксису могла стать взаимная конъюгация с образованием цитоплазматических мостиков и рекомбинацией геномной ДНК двух клеток (Gross, Bhattacharya, 2010). Нечто похожее на такое промежуточное звено, а именно конъюгация с образованием цитоплазматических мостиков, передачей геномной ДНК и с возможностью каждой клетки быть как донором, так и реципиентом, обнаружено у галофильных (солелюбивых) архей Haloferax (Halobacterium) volcanii (Rosenshine et al., 1989; Ortenberg et al., 1998).
Итак, половое размножение эукариот, возможно, является не просто аналогом, а прямым наследником прокариотического секса.
Плата за секс, или Двойная цена самцов
Мы выяснили, что секс для жизни исключительно полезен. Он залог стабильности в нашем нестабильном мире. Но, как известно, бесплатный сыр бывает только в мышеловке. Чем же живые существа платят за возможность быстро приспосабливаться?
Микробы, изредка меняющие несколько своих генов на заимствованные у соседей копии, возможно, платят за секс немного. Особенно если учесть, что механизм гомологичной рекомбинации, основанный на комплементарности, снижает риск попадания в геном чего-то совсем уж неподходящего, а возможность использовать чужую ДНК просто в качестве пищи служит дополнительным бонусом. Цена невысока, но и выигрыш от такого секса невелик. Он выше у амфиктических, раздельнополых организмов. Но и платят они за секс гораздо больше. Платить приходится за самцов, и цена получается удвоенная.
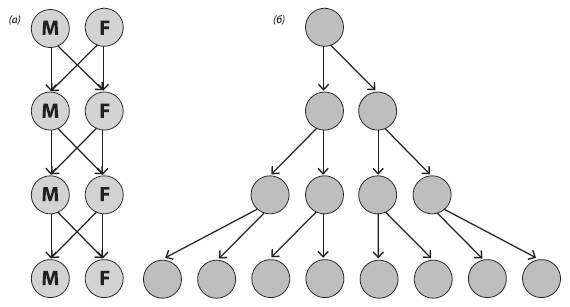
Суть в том, что при прочих равных бесполое клональное размножение (или самооплодотворение) ровно в два раза эффективнее, чем перекрестное оплодотворение с участием самцов (см. рисунок). Эту проблему рассмотрел выдающийся эволюционист Джон Мэйнард Смит (1920–2004) в книге The Evolution of Sex (1978).
Схема, иллюстрирующая «двойную цену самцов». У раздельнополых организмов половину потомства каждой самки составляют самцы, которые сами никакого потомства произвести не могут. При бесполом размножении все потомство состоит из самок (при самооплодотворении – из самостоятельно размножающихся гермафродитов). Поэтому при прочих равных размножение без участия самцов вдвое эффективнее, чем с самцами. На рисунке показана ситуация, когда каждая самка производит ровно двух потомков.
Получается, что самцы обходятся популяции непомерно дорого. Отказ от них дает значительный выигрыш в скорости размножения. Мы уже знаем, что переход от раздельнополости и перекрестного оплодотворения к бесполому размножению или самооплодотворению технически вполне возможен. Тому есть масса примеров как у растений, так и у животных. Однако бесполые расы и популяции самооплодотворяющихся гермафродитов почему-то до сих пор так и не вытеснили тех, кто размножается «обычным» образом, с участием самцов.
Из этого следует, что секс вообще (и раздельнополый секс в частности) должен давать настолько важные преимущества, что они перекрывают даже двойной проигрыш в эффективности размножения. Причем эти преимущества должны проявляться быстро, а не когда-нибудь через миллион лет. Повторим, что естественному отбору нет дела до отдаленных перспектив.
Еще о пользе секса, или Лучше меньше, да лучше
Рассмотрим пример, который показывает, что выигрыш от секса у раздельнополых организмов все равно перевешивает двойные потери в числе потомков. Такой пример нужно подбирать особенно тщательно, потому что нужен хороший контроль. В данном случае нужны две группы организмов (две популяции), одинаковые во всем, кроме одного – способности к сексу. И такие популяции биологам удалось создать.
Биологи из Орегонского университета (Morran et al., 2009) работали с уже знакомым нам червем C. elegans. Эти прекрасные животные словно нарочно созданы для проверки гипотез о пользе секса. Как мы помним, у них нет самок. Популяции состоят из самцов и гермафродитов, причем последних больше. Гермафродиты производят сперматозоиды и яйцеклетки и могут размножаться без посторонней помощи путем самооплодотворения. Самцы производят только сперматозоиды и могут оплодотворять гермафродитов. В результате самооплодотворения на свет появляются только гермафродиты. При перекрестном оплодотворении половина потомства оказывается гермафродитами, половина – самцами. Частота перекрестного оплодотворения в популяциях C. elegans обычно не превышает нескольких процентов. Чтобы определить эту частоту, не нужно следить за интимной жизнью червей – достаточно знать процент самцов в популяции.

У круглых червей Caenorhabditis elegans нет самок, а есть только самцы (вверху) и гермафродиты (внизу). Гермафродитов можно отличить по тоненькому длинному хвостику.
Следует пояснить, что самооплодотворение – не совсем то же самое, что бесполое (клональное) размножение, однако различия между ними быстро сходят на нет в череде самооплодотворяющихся поколений. После этого потомство перестает отличаться от родителей генетически, точно так же, как и при клональном размножении.
У C. elegans известны мутации, влияющие на частоту перекрестного оплодотворения. Одна из них, xol-1, смертельна для самцов и фактически приводит к тому, что в популяции остаются только гермафродиты. Другая, fog-2, лишает гермафродитов способности производить сперму и фактически превращает их в самок. Популяция, в которой все особи несут эту мутацию, становится обычной раздельнополой популяцией, как у большинства животных.
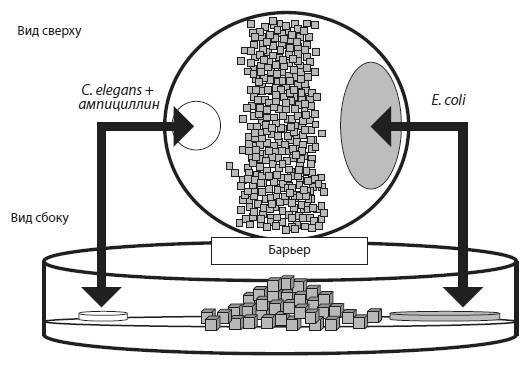
Схема экспериментальной установки. Молодых червей каждого нового поколения помещают в левую половину чашки (белый кружок). Чтобы добраться до еды (серый овал), они должны преодолеть барьер. Слабые особи, перегруженные вредными мутациями, не справляются с этой задачей. Из Morran et al., 2009.
Авторы при помощи классических методов (путем скрещиваний, а не генной инженерии) вывели две породы червей с почти одинаковыми геномами, различающимися только наличием мутаций xol-1 и fog-2. У первой породы была мутация xol-1, и нематоды размножались только самооплодотворением. У второй имелась мутация fog-2, так что эти черви размножались только путем перекрестного оплодотворения. К каждой породе прилагалась третья, лишенная обеих мутаций (дикий тип, ДТ). У ДТ частота перекрестного оплодотворения около 5 %. С этими тройками были проведены следующие две серии экспериментов.
В первой серии проверялась гипотеза о том, что перекрестное оплодотворение помогает избавляться от вредных мутаций. Эксперимент продолжался 50 поколений (червей, разумеется, а не экспериментаторов). Каждое поколение червей подвергалось действию химического мутагена – этилметансульфоната. Это увеличило частоту мутирования в четыре раза. Молодых животных помещали в чашку Петри, разделенную пополам барьером из крохотных кирпичиков (см. рисунок), причем червей сажали в одну половину чашки, а их пища (бактерии E. coli) находилась в другой половине. Таким образом, чтобы добраться до еды, а значит получить шанс выжить и оставить потомство, черви должны были преодолеть барьер. Тем самым экспериментаторы повысили эффективность очищающего отбора, который отсеивает вредные мутации. В обычных лабораторных условиях эффективность отбора низка, потому что черви окружены пищей со всех сторон. Это позволяет выжить даже очень слабым животным, перегруженным вредными мутациями. В новой экспериментальной установке этой уравниловке был положен конец. Чтобы преодолеть стенку, червь должен быть здоров и крепок.