Третий белок отличался от сестринских белков сильнее, чем они друг от друга. Он выполнял роль «внешней группы», позволяя понять, какие аминокислотные замены произошли в первом, а какие во втором сестринском белке. Например, если у внешнего белка в данной позиции стоит аминокислота K (лизин) и такая же аминокислота стоит здесь у первого из двух сестринских белков, а у второго в этом месте находится другая аминокислота (например, Y – тирозин), то считалось, что K в данной позиции – это «предковое», исходное состояние, а у второго сестринского белка произошла замена K на Y.
К этим трем белкам добавлялся четвертый, еще более далекий от сестринских, чем третий. Если у четвертого белка в данной позиции стоит аминокислота K (как на рисунке), то делался вывод, что у второго сестринского белка произошло эволюционное изменение, увеличившее дистанцию между белками 2 и 4: белки разошлись. Если бы у четвертого белка здесь стояла аминокислота Y, то был бы сделан вывод, что в результате изменения, затронувшего белок 2, белки 2 и 4 сблизились.
В общей сложности было обработано 13,6 млн таких четверок. Для каждой четверки определялось число замен, ведущих к сближению последовательностей и к их расхождению. По соотношению этих двух величин можно понять общую тенденцию: расходятся ли белки, сближаются или балансируют вокруг некого постоянного уровня сходства последовательностей.
Оказалось, что даже у наиболее удаленных друг от друга гомологичных белков тенденция к расхождению значительно преобладает над тенденцией к сближению. Следовательно, «вселенная древних белков» продолжает расширяться и пределы областей, соответствующих их функциям, за 3,5 млрд лет так и не были достигнуты. Эволюция этих белков была крайне медленной. Что же ее сдерживало? Решению этой проблемы посвящена вторая часть статьи, приводящая на память другой рассказ Борхеса – «Сад расходящихся тропок».
Лабиринт расходящихся тропок
Обычного отрицательного отбора, отсеивающего вредные мутации, явно недостаточно для того, чтобы объяснить медленную эволюцию древних белков. Предположение о том, что каждая аминокислота, стоящая в данной позиции, влияет на приспособленность одинаковым образом независимо от «контекста» (т. е. от того, какие аминокислоты стоят в других позициях в том же белке или в других белках, взаимодействующих с ним), соответствует представлению о легкопроходимом ландшафте приспособленности (см. рисунок ниже). Но со времен LUCA в каждом синонимичном сайте (так называют те нуклеотиды в цепи ДНК, изменение которых не влияет на структуру кодируемого белка) произошло уже свыше 100 замен. Синонимичные нуклеотидные замены – это пример движения по ровным горизонтальным поверхностям ландшафта приспособленности. Из этого следует, что в случае хорошей проходимости ландшафта изучаемые белки давным-давно должны были освоить всю потенциально доступную им область в пространстве последовательностей, и тогда мы не наблюдали бы их продолжающегося расхождения.
Резко уменьшить проходимость ландшафта приспособленности – превратить его в лабиринт узких тропок или горных хребтов – может эпистаз. Так называют взаимное влияние разных мутаций (аллелей) на вредность или полезность друг друга. Замена одной аминокислоты может менять эффективность работы белка не сама по себе, а в зависимости от комбинации других аминокислот, занимающих определенные позиции в том же белке. Также эффект мутации может зависеть от других белков, находящихся в непосредственном взаимодействии с белком-мутантом. Иными словами, для определения смысла биологического текста важен контекст. В этом и состоит суть эпистаза. Мы еще не раз встретимся с этим явлением в последующих главах.
Наличие эпистаза предполагает, что многие аминокислотные замены являются допустимыми только в определенном окружении. Если окружение (контекст) неподходящее, то данная замена будет снижать приспособленность, и отбор ее выметет. Это явление в эволюции белков изучалось ранее на конкретных примерах[22]. Если эпистаз широко распространен, белок может добраться до многих потенциально достижимых пунктов в пространстве последовательностей только долгим обходным путем.
Для проверки гипотезы о действенности эпистаза в эволюции белков Поволоцкая и Кондрашов придумали хитроумный тест, основанный на сопоставлении темпов накопления «сближающих» и «разводящих» мутаций с дистанциями между белками. Основная идея состояла в том, что гипотезы о наличии и отсутствии эпистаза дают противоположные предсказания о частоте закрепления сближающих и разводящих мутаций. Если влияние эпистаза невелико, то у двух близких белков темп накопления разводящих мутаций должен быть высоким, а по мере расхождения последовательностей он должен снижаться (если ландшафт приспособленности легкопроходимый, то белки сначала свободно и быстро «блуждают» по ровному плато, соответствующему данной функции, уходя все дальше от исходной точки, но по мере приближения к его краям их расхождение замедляется). Темп накопления сближающих мутаций при этом должен оставаться примерно постоянным. Если же эпистаз оказывает сильное влияние на эволюцию белков, все должно быть наоборот: разводящие мутации должны накапливаться с примерно постоянной скоростью, а темп накопления сближающих мутаций по мере расхождения белков должен снижаться.
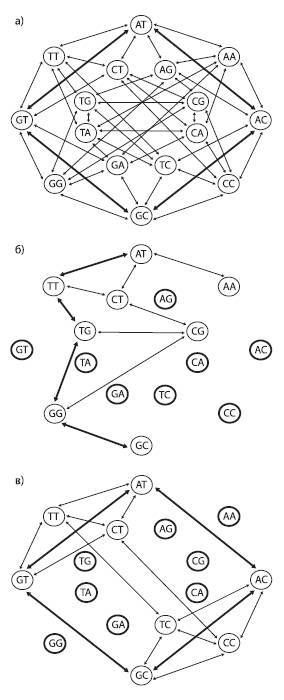
Пространство последовательностей можно изобразить в графа, вершины которого соответствуют разным последовательностям (в данном случае показаны последовательности из двух нуклеотидов), а ребра – единичным эволюционным событиям (нуклеотидным заменам). В первом случае (a) ландшафт приспособленности представляет собой сплошное ровное плато: все 16 последовательностей имеют одинаково высокую приспособленность и все возможные мутации разрешены (не будут отбракованы отбором). Два кратчайших пути, соединяющие последовательности AT и GC, состоят всего из двух мутационных шагов (жирные стрелки). Во втором случае (б) половина последовательностей имеют пониженную приспособленность (отмечены жирными кругами). При этом снижение приспособленности определяется не конкретным нуклеотидом в конкретной позиции, а уникальной комбинацией обоих нуклеотидов (т. е. имеется эпистаз). Это резко снижает проходимость ландшафта: число доступных траекторий уменьшается и кратчайший путь между двумя пунктами (например, от AT к GC) удлиняется. В третьем случае (в) тоже половина последовательностей имеют пониженную приспособленность, но эпистаза нет: нуклеотиды A и G во второй позиции снижают приспособленность независимо от состояния первой позиции. Отсутствие эпистаза способствует тому, что ландшафт остается легкопроходимым, и от AT к GC можно прийти всего за два шага, как и в первом случае. По рисунку из Povolotskaya, Kondrashov, 2010.
Проведенные расчеты подтвердили гипотезу о сильном влиянии эпистаза на эволюцию белков. Оказалось, что темп накопления разводящих мутаций не зависит от дистанции между белками. В каждый момент времени лишь около 2 % аминокислотных позиций могут быть изменены без снижения приспособленности, хотя в долгосрочной перспективе более 90 % позиций могут измениться – но к этим изменениям нужно идти долгими обходными путями по лабиринту узких «горных хребтов» ландшафта приспособленности. У близких, недавно разошедшихся белков темп накопления сближающих мутаций очень высок, потому что у близких белков аминокислотный «контекст» для каждой позиции является сходным. Поэтому та аминокислота, которая недавно стояла в данной позиции, с большой вероятностью может «вернуться» на свое место, и это не снизит приспособленность. Напротив, у сильно различающихся белков данная позиция уже находится в разных контекстах, и поэтому «возвращение» аминокислоты, которая стояла здесь у далекого предка, скорее всего, снизит приспособленность, и мутация будет отсеяна.