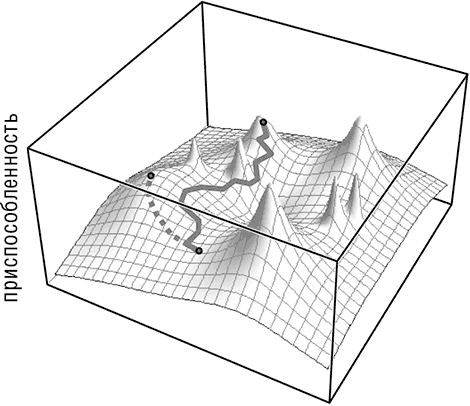
Рис. 1–2. Эволюционные траектории на неровном адаптивном ландшафте. Пунктирной линией обозначается эволюционная траектория при высоком значении эффективного размера популяции. Сплошной линией обозначается эволюционная траектория при низком значении эффективного размера популяции.
Джон Мейнард Смит и, позднее, Джон Гиллеспи разработали теорию и компьютерные модели для демонстрации существования особого режима нейтральной эволюции, который слабо зависит от эффективного размера популяции и актуален даже в популяции бесконечного размера с сильным отбором. Этот способ нейтрального закрепления мутаций стал известен как «генетическая тяга» и относится к ситуациям, в которых одна или несколько нейтральных или даже умеренно вредных мутаций распространяются в популяции и в конечном итоге закрепляются, будучи связанными с полезной мутацией. Иными словами, нейтральные или вредные аллели «двигаются в одной повозке» вместе с полезным аллелем (Barton, 2000). Похоже, что некоторые данные и модели популяционной генетики свидетельствуют, что «движение в одной повозке» даже важнее для эволюции популяции с половым размножением, чем дрейф. Очевидно, что эффект «езды в одной повозке» обусловлен совокупным воздействием естественного отбора и нейтральной изменчивостью в различных участках генома и, в отличие от дрейфа, может происходить даже в популяции бесконечно большого эффективного размера (Gillespie, 2000).
За счет эффекта «движения в одной повозке» даже в больших популяциях могут закрепляться умеренно вредные мутации, что, соответственно, дает этой популяции возможность пересекать долины адаптивного ландшафта.
Положительный и очищающий (отрицательный) отбор: классификация форм отбора
Дарвин думал о естественном отборе в первую очередь с точки зрения закрепления благоприятных изменений. Он понимал, что эволюцией отсеиваются вредные изменения, но не интерпретировал эту ликвидацию в одной плоскости с естественным отбором. С развитием СТЭ понятие отбора было расширено за счет включения «очищающего» (отрицательного) отбора, который в некоторых фазах эволюции оказывается более распространенным (на самом деле на порядок более распространенным), чем «дарвиновский» положительный отбор. По сути, очищающий отбор – это просто элиминация неприспособленных особей. Тем не менее выделение этого процесса в особую форму отбора представляется оправданным и важным, потому что оно подчеркивает ключевую роль элиминации в формировании (сдерживании) биологического разнообразия на всех уровнях. Проще говоря, изменение допускается, только если оно не наносит существенного вреда никаким из выживающих особей. Интересен и открыт вопрос, до какой степени эти ограничения фактически сужают пространство, доступное для эволюции, и я коснусь этого вопроса позднее (см., в частности, гл. 3, 8 и 9).
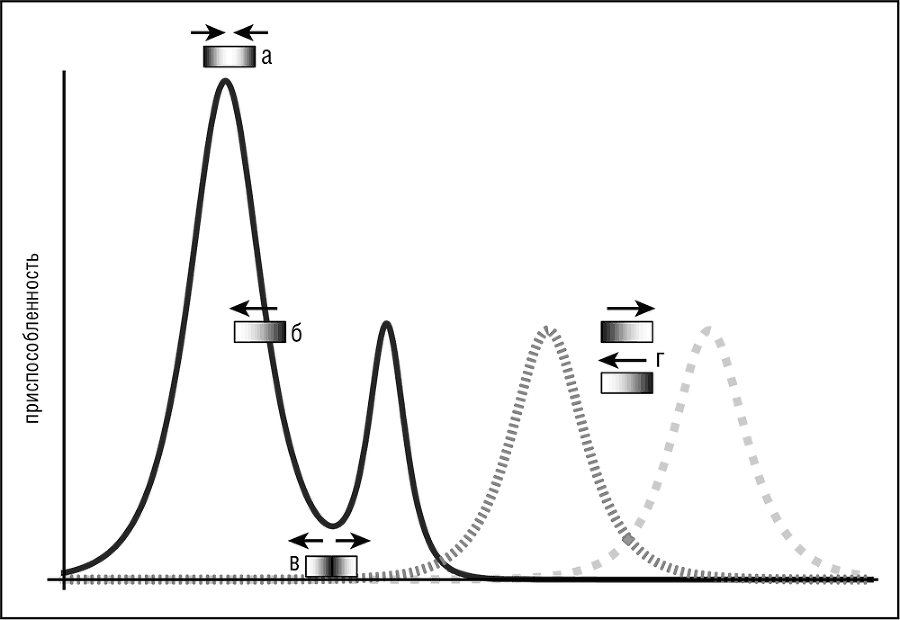
Тонкая, но важная разница существует между очищающим отбором и стабилизирующим отбором, который является еще одной из форм отбора, которая действует на распределение частот отличительных признаков. Таким образом, выделяются следующие формы отбора: стабилизирующий отбор, основанный в первую очередь на очищающем отборе, движущий отбор, обусловленный положительным (дарвиновским) отбором, и более экзотические режимы дизруптивного и балансирующего отбора, которые являются результатом сочетания многочисленных ограничений (см. рис. 1–3).
Рис. 1–3. Четыре различные формы отбора в эволюционирующей популяции: а — стабилизирующий отбор (адаптивный ландшафт представлен сплошной линией); б — движущий отбор (адаптивный ландшафт представлен сплошной линией); в — дизруптивный отбор (адаптивный ландшафт представлен сплошной линией); г — балансирующий отбор (адаптивный ландшафт периодически меняется, переключаясь между двумя пунктирными линиями)
Синтетическая теория эволюции
Объединение дарвиновской теории эволюции и генетики, состоявшееся в основополагающих исследованиях Фишера, Райта и Холдейна, подготовило почву для рождения синтетической теории эволюционной биологии. Само название идет от одноименной книги, опубликованной Джулианом Хаксли в 1942 году (Huxley, 2010), однако концептуальная структура СТЭ полностью сформировалась только в 1959 году в ходе мероприятий, посвященных 100-летнему юбилею «Происхождения…». Новая синтетическая теория стала результатом работы многих выдающихся ученых. Можно утверждать, что главными архитекторами СТЭ были экспериментальный генетик Феодосий Добржанский, зоолог Эрнст Майр и палеонтолог Джордж Гейлорд Симпсон. Экспериментальные и полевые работы Добржанского с плодовой мушкой Drosophila melanogaster принесли насущные фактические свидетельства в поддержку теории популяционной генетики и стали первой крупномасштабной экспериментальной проверкой идеи естественного отбора. Книга Добржанского «Генетика и происхождение видов» (Dobzhansky, 1951) явилась основным программным документом СТЭ, в котором он сузил понятие эволюции до «изменения частоты аллеля в генетическом пуле». Знаменита также крылатая фраза Добржанского о том, что «ничто в биологии не имеет смысла, кроме как в свете эволюции»[13] (см. больше о «смысле» в прил. I). Заслугой Эрнста Майра, как никакого другого ученого, является серьезная, крайне влиятельная попытка теоретического решения принципиальной проблемы, поставленной Дарвином, – происхождения видов. Майр сформулировал так называемую биологическую концепцию вида, согласно которой видообразование происходит, когда две популяции (размножающиеся половым путем) изолированы друг от друга достаточно долго, чтобы обеспечить необратимую генетическую несовместимость (Mayr, 1963).
Симпсон реконструировал наиболее полную (на тот момент) картину эволюции жизни на основании палеонтологической летописи (Simpson, 1983). Замечательно, что Симпсон осознал стазис (отсутствие существенных изменений) в эволюции большинства видов и резкую смену доминантных видов. Он ввел понятие квантовой эволюции, которое предвосхитило теорию прерывистого равновесия, предложенную Стивеном Джеем Гулдом и Нильсом Элдриджем (см. гл. 2).
Консолидация СТЭ в 1950-х годах была довольно странным процессом, сопровождавшимся странным «затвердеванием» (выражение Гулда) основных идей Дарвина (Gould, 2002). Так, доктрина СТЭ фактически отбросила идею Райта о случайном дрейфе генов и его эволюционной важности и стала бескомпромиссно панадаптационистской. Более того, сам Симпсон отказался от идеи квантовой эволюции, так что градуализм продолжал оставаться одним из неоспоримых столпов СТЭ. Такое «затвердевание» сделало СТЭ относительно узкой, в некотором смысле даже догматичной, системой.
Чтобы продолжить обсуждение эволюции эволюционной биологии и ее преображение в век геномики, представляется необходимым кратко резюмировать основные принципы эволюции, впервые сформулированные Дарвином, затем усовершенствованные первым поколением биологов-эволюционистов и, наконец, кодифицированные в СТЭ. Мы будем возвращаться к каждому из этих ключевых моментов на протяжении всей книги.
1. Ненаправленное случайное изменение – это главный процесс, обеспечивающий материал для эволюции. Дарвин впервые показал, что случайность является основным фактором в истории жизни, и это, несомненно, было одной из его наиболее важных идей. Дарвин также признавал роль направленной, ламарковской изменчивости и в последующих изданиях «Происхождения…» склонялся даже к более весомой роли этого механизма эволюции. Однако СТЭ твердо настаивает на том, что случайные мутации являются единственным источником эволюционно значимой изменчивости.
2. Действие эволюции заключается в фиксации редких выгодных изменений и элиминации вредных изменений. Согласно Дарвину и СТЭ, в этом состоит процесс естественного отбора, который, наряду со случайной изменчивостью, является основной движущей силой эволюции. Естественный отбор, очевидно сходный и навеянный «невидимой рукой» рынка, которая, по теории Адама Смита, управляет экономикой, был первым из когда-либо предложенных механизмов эволюции, который был прост и правдоподобен и не требовал изначально мистического подхода. Таким образом, это вторая ключевая идея Дарвина. Сьюэл Райт подчеркивал, что случайность может играть вспомогательную роль не только в возникновении, но также и в закреплении изменений в ходе эволюции с помощью дрейфа генов, в результате которого случайно сохраняются нейтральные или умеренно вредные изменения. Согласно теории популяционной генетики, дрейф генов особенно значим в небольших популяциях, проходящих через «бутылочное горлышко». «Генетическая тяга», или «езда в одной повозке», – это другая форма случайного закрепления невыгодных мутаций. Однако СТЭ в ее догматизированной форме фактически отрицает стохастические процессы в эволюции, кроме возникновения изменений, и придерживается полностью адаптационистского (панадаптационистского) взгляда на эволюцию. Такая модель неизбежно приводит к концепции «прогресса», постепенного улучшения «органов» в ходе эволюции. Дарвин поддерживал эту идею как основное направление развития, несмотря на четкое понимание, что организмы все еще далеки от совершенства в плане адаптивности, как можно прекрасно увидеть на примере рудиментарных органов, и несмотря на свое резко отрицательное отношение к любым формам ламарковского внутреннего стремления к совершенству. СТЭ уходит от прогресса как антропоморфной идеи, но тем не менее поддерживает общую концепцию эволюции от простых форм к сложным.