Пандативы, экзаптация, эволюция как ремесленник и ошибочность панглоссианской парадигмы эволюции
Пусть и неявно, но принципу градуализма был брошен вызов гипотезой Оно об эволюции генов и геномов путем дупликации, a затем, в явной форме, концепцией прерывистого равновесия. Адаптационистская программа эволюционной биологии подверглась решительной, сметающей все на своем пути атаке в статье 1979 года «Пандативы Святого Марка» Гулда и Левонтина (Gould and Lewontin, 1979), одной из самых необычных и влиятельных статей в истории биологии. Гулд и Левонтин саркастически описали адаптационистскую картину мира как панглоссианскую парадигму, названную так в честь примечательного персонажа вольтеровского «Кандида», который утверждал, что «все к лучшему в этом мире» [пер. Ф. Сологуба] (даже катастрофы). Гулд и Левонтин подчеркивали, что вместо того, чтобы стряпать на скорую руку «сказки просто так»[25] о правдоподобных адаптациях, эволюционным биологам следовало бы искать объяснение наблюдаемых черт организации биологических организмов исходя из плюралистского подхода, который принимает во внимание не только отбор, но также и внутренние ограничения, случайный дрейф и другие факторы. Метафора пандатива означает, что многие функционально важные элементы биологической организации вовсе не эволюционировали как специальные устройства для выполнения определенных функций, но скорее являются продуктами неадаптивных архитектурных ограничений, подобно пандативам (spandrels), появляющимся в арках соборов и других зданий исключительно вследствие требований конструкции, и могут использоваться для различных целей, например для украшения собора (см. рис. 2–2). Процессу использования пандативов в биологии было дано специальное название экзаптация, и Гулд провозгласил его важным путем эволюции (Gould, 1997a). Концепция пандативов связана с почти нейтральной теорией, но в каком-то смысле идет дальше и подходит ближе к сути эволюционного мышления, показывая, что даже те фенотипические черты, которые выглядят как типичные адаптации, не обязательно эволюционировали под давлением естественного отбора.

Рис. 2–2. Один из пандативов базилики Святого Марка в Венеции. Фото Марии Шнитцмейер, Викисклад.
В более ранней статье по сходной тематике Франсуа Жакоб (один из первооткрывателей регуляции генов и автор нескольких других плодотворных идей в бактериальной генетике, см. гл. 5) ввел метафору мастера-самоучки. Отталкиваясь прежде всего от сравнительного анализа механизмов развития, Жакоб положил в основу своих рассуждений, что эволюция действует не как инженер или дизайнер, а скорее как ремесленник-самоучка, причем чрезвычайно зависимый от предыдущего опыта при решении стоящих перед ним проблем: «Сложно проводить аналогии между естественным отбором и какими-то аспектами человеческого поведения. Однако если очень хочется поиграть в сравнения, то можно сказать, что естественный отбор работает не как изобретатель или инженер. Он работает как дилетант – мастер на все руки, который не знает точно, что он собирается создать, и при этом использует все, что подвернется под руку, будь то обрывки ниток, куски дерева или старые коробки; короче, он действует как тот мастеровой, который использует все, что есть в его распоряжении, чтобы сделать хоть что-то, лишь бы работало» (Jacob, 1977).
Ключевым выводом концепции ремесленника-самоучки становится то, что итоговый результат эволюции непредсказуем, или по крайней мере его невозможно предсказать, не зная в деталях всех предшествующих событий. Другими словами, если взять и «проиграть эволюционную пластинку заново» (любимая метафора Гулда) в некоем мысленном эксперименте, то результат будет отличен от того, что мы наблюдаем в реальности, возможно до неузнаваемости; мы вернемся к этому обсуждению позднее в этой книге (см. гл. 13).
Эволюция в мире микробов и вирусов и трехдоменное древо жизни
Вероятно, в ходе развития биологии наибольшее влияние на изменение представления об эволюции оказало распространение эволюционных исследований на мир микробов, а именно одноклеточных эукариот (протист), прокариот (бактерий и архей) и вирусов. Дарвиновское представление об эволюции и все достижения эволюционной биологии нескольких последующих десятилетий базировались исключительно на исследовании животных и растений, тогда как одноклеточные эукариоты (протисты) и бактерии (монеры) были сугубо номинально размещены у корня древа жизни Эрнстом Геккелем и его последователями. Хотя к 1950-м годам генетический анализ бактериофагов и бактерий продвинулся настолько, что стало очевидным, что эти формы жизни обладают эволюционирующими геномами, СТЭ не принимала во внимание эти открытия. То, что бактерии (не говоря уже о вирусах) эволюционируют по тому же самому принципу и с использованием тех же механизмов, что и животные и растения, отнюдь не очевидно, учитывая все их разительные биологические отличия от многоклеточных организмов, и в особенности из-за отсутствия у них типичного полового размножения и репродуктивной изоляции, ключевых для видообразования среди животных и растений.
Фактически прокариоты стали «видны» эволюционным биологам в 1977 году, после выхода революционной работы Вёзе и его коллег по филогенезу рРНК (Woese, 1987)[26]. Рассмотренное в общем контексте, открытие Вёзе является эпохально важным и, возможно, даже заслуживает сравнения с открытием структуры ДНК. Вёзе установил, что в одной молекулярной структуре, а именно последовательности нуклеотидов рРНК, выявляется очевидная консервативность во всем диапазоне клеточных форм жизни. Кроме того, чрезвычайно информативным оказался и филогенетический анализ этой универсальной консервативной молекулы: он показал, что рРНК, в некотором приближении, эволюционирует с постоянной скоростью, то есть подчиняется модели молекулярных часов. Это привело к еще одному важному открытию, ставшему одним из символов эволюционной биологии конца ХХ века, – трехдоменному древу жизни (см. рис. 2–3; Woese et al., 1990). Тремя доменами являются бактерии, археи и эукариоты. Домен архей был открыт Джорджем Фоксом и Вёзе сравнительным анализом рРНК, когда в новой группе ничем, казалось бы, не примечательных «бактерий» обнаружились существенные отличия как от остальных бактерий, так и от более сложных эукариотических организмов. В дополнение к разграничению трех доменов, Вёзе и его коллеги использовали филогенетический анализ рРНК для идентификации нескольких основных ветвей архей и бактерий (Woese, 1987). Из этого следовало, что эволюция прокариот столь же доступна для изучения, как эволюция сложных эукариот, – концепция, чуждая микробиологам до работы Вёзе (Stanier and Van Niel, 1962). Благодаря достижениям Вёзе, его сотрудников и последователей появилась все усиливающаяся тенденция приравнивать филогенетическое древо рРНК, с его трехдоменной структурой, к древу жизни Дарвина и Геккеля (Pace, 2009а, 2006). В течение нескольких лет после публикации открытий Вёзе стало ясно, что топологически древо рРНК (по крайней мере, в своих основных чертах) конгруэнтно деревьям некоторых из самых консервативных белков, таких как рибосомные белки, факторы трансляции, субъ единицы ДНК-зависимой РНК-полимеразы и мембранные АТФазы.
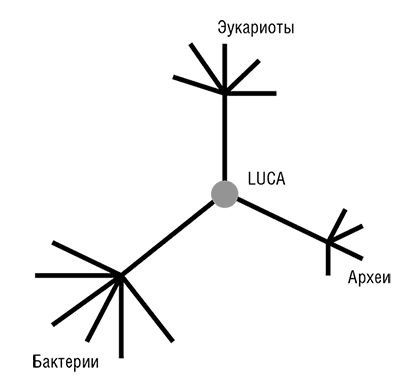
Рис. 2–3. Трехдоменное древо жизни Вёзе.
Две группы исследователей независимо друг от друга пришли к блестящей идее о том, как определить положение корня в эволюционном дереве, которое до этого было бескорневым (рис. 2–3). Для этой цели можно использовать древние паралоги, которые представлены в (почти) всех организмах и, таким образом, можно с уверенностью заключить, возникли в результате дупликации, предшествующей последнему общему предку всех живых организмов (LUCA). Когда дерево строится совместно для двух паралогичных множеств древних ортологов, положение корня между ними определено однозначно, и таким образом корень может быть выведен для каждого из множеств ортологов (см. рис. 2–4; Gogarten et al., 1989; Iwabe et al., 1989). Результаты анализа двух пар древних паралогов, факторов трансляции и субъединиц мембранных АТФаз были полностью совместимы и поместили корень на бактериальную ветвь, установив таким образом кладу архей-эукариотов (см. рис. 2–4). Тем не менее даже в догеномную эпоху было ясно, что не все деревья белок-кодирующих генов имеют ту же топологию, что и дерево рРНК; причины этих отличий оставались неясными и, как предполагалось, включали (за исключением артефактов метода) горизонтальный перенос генов (ГПГ. Smith et al., 1992). Эти расхождения оставались лишь интересным дополнением к трехдоменному ДЖ, но все резко изменилось с наступлением эры геномики.