AL алтайцы, DR индусы, дравиды, ES эскимосы, GE грузины и армяне, GM немцы, HA китайцы хань, IB Iberian peninsula, IS Исландцы, IN индусы, индо-арии, population, IT итальянцы, KG киргизы, KT казанские татары, KZ казахи, MA арабы Среднего Востока, MO монголы, MY малазийцы, NE ненцы, NW норвежцы, PE персы (Иран), RU русские, SA саамы, SC шотландцы, SL селькупы, TB тибетцы, TU турки, UG уйгуры, UZ узбеки.
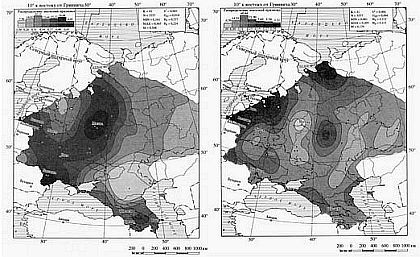
В русском генофонде почти половина Y хромосом относится к гаплогруппе R1a. Основной ареал этой гаплогруппы — Восточная Европа, в центре которой у русских максимальные частоты R1a (до 62 % в Белгородской и Орловской областях). Частота данной гаплогруппы высока также у белорусов. Будем называть ее русской.
Типично западноевропейская гаплогруппа R1b у русских встречается в 6 % случаев, у украинцев — 6 %, у белорусов — 3 %. Мы можем считать ее доарийской.
Геногеографические карты распределения частот гаплогрупп Y-хромосомы: R1a и R1b. Внизу показаны те же рисунки с затемнением зон с малыми значениями частот.
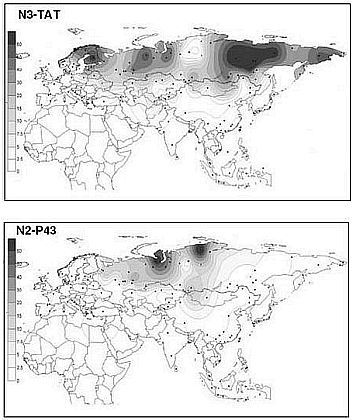
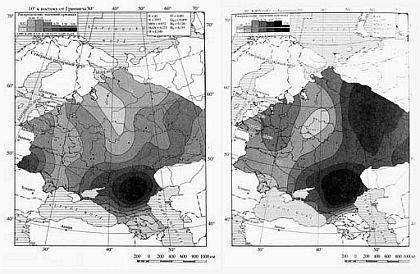
Гаплогруппа N3 в русском генофонде присутствует в 18 % Y хромосом, а у белорусов и украинцев низка. Ее основной ареал — Сибирь и Восточная Европа (в Сибири до 90 % в отдельных популяциях). По другим признакам генофонды Восточной Европы и Сибири резко различны. Возможно, эта гаплогруппа отражает династии, протянувшиеся от первых ариев южносибирских степей. Значительно менее распространенной среди русских (2 %) и имеющей ядро в Западной Сибири является гаплогруппа N2. У русских она встречается только на севере ареала. Предположительно будем считать данные гаплогруппы раннеарийскими.
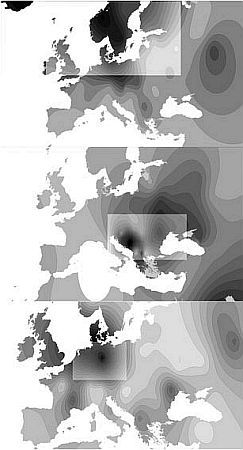
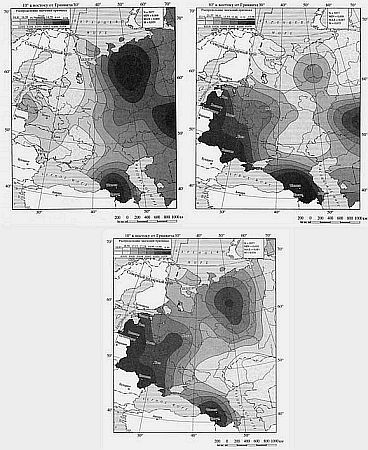
Из гаплогрупп I1a, I1b и I1с первая максимально распространена в Скандинавии (25–40 %), вторая — на Балканах, третья — в северной Скандинавии. У русских доминирует «балканская» гаплогруппа вдвое преобладающая над «скандинавской». У белорусов соотношение в пользу «балканского» варианта 15:1, у украинцев 5:1. Логично считать ее позднеарийской — связанной с двумя ветвями арийской миграции.
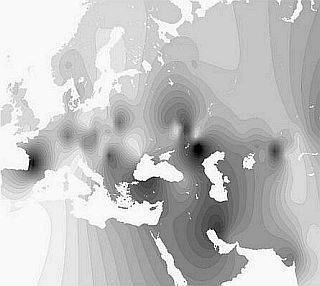
Гаплогруппу J2, которая преобладает на Ближнем Востоке и на Кавказе, встречается в Южной Европе, а в русском ареале распространена слабо (2 %), можем назвать кавказоидной.
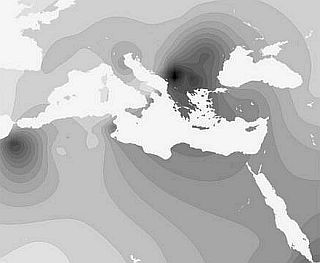
Гаплогруппа e3b является типично средиземноморской или североафриканской (частота в Северной Африке — до 80 %), откуда она распространилась в Средиземноморье, в наибольшей степени — на Балканах (10–20 %). В русском ареале она малозаметна (2 %).
Геногеографические карты распределения частот гаплогрупп Y-хромосомы: N3a и N2.
Геногеографические карты распределения частот гаплогрупп Y-хромосомы: I1a, I1b, I1c. Продвижение очагов образования родовых линий происходит с юга на север, что может быть связано с отступлением ледника. В осветленных окнах показаны зоны с высокими значениями частот.
Геногеографические карты распределения частот гаплогрупп J2 Y-хромосомы
Геногеографические карты распределения частот гаплогрупп е3b1 Y-хромосомы.
Доводы о родстве современных греков со славянами развеиваются тем фактом, что среди славян распространены высокие частоты гаплотипов R1a и I1b и низкие частоты средиземноморского гаплотипа е3b1. Среди греков общая частота гаплотипов R1a и I — приблизительно 25 %, а гаплотипа е3b1 достигает почти 20 %. На Пелопоннесе гаплотип е3b1 достигает частоты 47 %.
Дифференциация по локусу DM является еще одним расово-диагностическим методом.
При миотонической дистрофии наблюдается увеличение повторов триплета в гене хромосомы 19 до сотен и тысяч. У здоровых людей триплетный локус DM имеет от 5 до 30 или несколько более повторов. Группа аллелей с числом повторов 11–14 варьирует у разных народов в пределах 10–15 %. Между 5 и 11-членными и 15-19-членными повторами встречаемость аллелей крайне низка. 20–22 повтора составляют третью, наиболее слабую волну встречаемости, а за ней — область больших значений повторов с размытым спадом частоты встречаемости.
Локус DM ведет себя как нейтральный маркер, находясь в некодирующей части ДНК. У разных популяций частота встречаемости повторов той или иной длины оказывается различной. Например, у русских пятичленный повтор наиболее распространен, у славян он достигает 40–50 %.
У башкир встречаемость пятичленного повтора значительно ниже, а следующим по встречаемости остается 12-13-кратный повтор, как и у русских, но его частота заметно увеличивается, а также сравнимой становится частота встречаемости 11 кратного повтора. Кроме того, заметно возрастает значение 20-кратного повтора. У якутов 5-кратный повтор еще ниже и уступает заметно более распространенным 13-14-кратным повторам.
Распространенность 5-членного локуса выделяет в географическом пространстве великорусский регион с центром в Москве. Напротив, 11-членный локус дает вокруг Москвы низкую встречаемость. 12-ти и 13-ти членные локусы наиболее распространены в пространстве между Каспийским и Черным морями, фиксируя альтернативу генетической особенности великорусского ядра, отмеченного 5-членным локусом. Карта гетерозиготности локуса DM по всем аллелям демонстрирует совпадение тенденций и локальных максимумов и минимумов с картами по каждому локусу, что говорит о высоком уровне корреляции. Высокая частота аллеля снижает число гетерозигот; поэтому локальные максимумы на картах отдельных локусов могут совпадать с локальными минимумами на картах гетерозиготности. Карта гетерозиготности по DM показывает особую выделенность 5-кратного повтора в сравнении со всеми остальными вариантами кратности — карта 5-кратного повтора дает основной вклад в общую изменчивость генофонда, с высокой точностью повторяя карту гетерозиготности.
На карте первой главной компоненты изменчивости локуса DM московский регион выделяется как локальный минимум, зона между северным Причерноморьем и Прикаспием — как локальный максимум. Еще одна зона максимума изменчивости возникает вокруг Казани и севернее. На Кавказе возникает еще один резкий минимум, демонстрирующий географическую зависимость признака, имеющую место наряду с очевидной расово-этнической зависимостью в восточно-европейском пространстве.
Микросателлитные локусы (DM и другие) дают в руки ученым широкий набор инструментов для расовой диагностики. Интегральный анализ большого набора микросателитных локусов (частот их повторов) позволяет с высокой вероятностью определять расовую принадлежность индивида, а в масштабных исследованиях даже уточнить расовые разграничения, которые зачастую опираются на зыбкую основу лингвистических и физиономических наблюдений.
В целом 5-членный локус DM выделяет среднерусское население среди всего остального населения восточно-европейского пространства и фиксирует его генетическую особенность.
Микросателлитные локусы DM с числом повторов 5, 11 и 12-кратные повторы и первая главная компонента изменчивости локуса DM
[Лимборская С.А. и др. Этногеномика и геногеография народов Восточной Европы. — М.: Наука, 2002.]
Карты генетических расстояний от средних частот популяций индоевропейской, уральской и альпийской языковых семей
[Лимборская С.А. и др. Этногеномика и геногеография народов Восточной Европы. — М.: Наука, 2002.]
Диагностирует русский генофонд мутация, противодействующая заражению вирусом иммунодефицита. В целом русский ареал повторяет максимум частоты в беломоро-балтийской зоне и спад частоты к югу. Но в отличие от глобального тренда частота соответствующей мутации падает более решительно не к востоку, а к западу от зоны максимальных частот. Псковские популяции, примыкающие к этой зоне, имеют заметно более низкие частоты мутации (5–9%) в сравнении с центральными и даже восточными русскими популяциями (10–15 %). Также аномально низкую частоту имеет вологодская популяция (5 %) в сравнении с расположенным рядом русским максимумом (16–19 %). Локализация максимальных частот также задает исследователям загадки — изолированные друг от друга популяции дают резкое отклонение в сторону увеличения от среднерусской частоты примерно на одну и ту же величину.