Фундаментальный вывод, следующий из геногеографии, свидетельствует, что классификация по признакам европеоидности-монголоидности, принятая антропологами для русского и славянского населения, является совершенно негодной. «Тени» монголоидности — скорее штамп научного языка, чем реальность. Русский генотип, как видно из его пространственного распределения, вполне допускает то, что называют «налетом монголоидности», но на самом деле является лишь внутрирусской расовой вариацией.
Второй фундаментальный вывод состоит в том, что зона западной и северной Украины, юга Белоруссии и примыкающая к ним территория южнее Воронежа — генетически проблемна. При том что эта зона представлена единым антропологическим типом и признаки в ней понтийской расы, более проявленные южнее, еще малозаметны, она отделена как от собственно русского пространства, так и от европейского антропологического континента. К сожалению, загадочный район Ровно-Житомир остался практически не изученным антропологами. Внимательное отношение к нему, между тем, диктуется особенностями, которые этот район демонстрирует во всех представлениях геногеографического материала (то есть, по разным вариантам выборки групп генов — по трем первым главным компонентам или по генетическим «расстояниям» от некоей средней нормы).
Третий вывод (в котором мы, наконец, совпадаем с авторами исследования «Восточные славяне») состоит в явном указании на неевропейское происхождение генотипов не только Поволжья, Приуралья и Казахстана, но и Кавказа и Малой Азии, и, что самое любопытное, Прибалтики, Карелии и т. н. Фенно-Скандии. При этом речь идет не о признаках монголоидности, а обо всем комплексе генетических признаков — интегральном генетическом различии.
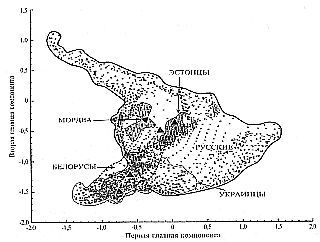
Общность генетики Русского мира неплохо прослеживается в координатах главных компонент, когда в них выстраиваются «этнические облака» — группы точек, каждая из которых отражает средний показатель замера в отдельной популяции.
Об общности или родственности генофонда говорит тот факт, что в пространстве первой-второй, второй-третьей и первой-третьей главных компонент, рассчитанных из анализа генетического разнообразия народов Восточной Европы, изменчивость популяций эстонцев и мордвы полностью поглощается изменчивостью великороссов. Причем обе популяции оказываются чрезвычайно близки, несмотря на свое географическое и историческое разобщение.
Изменчивость генофонда Восточной Европы в пространстве главных компонент (треугольниками обозначены центры тяжести «этнических облаков»).
[Лимборская С.А. и др. Этногеномика и геногеография народов Восточной Европы. — М.: Наука, 2002.]
В целом великорусское «этническое облако» в пространстве главных компонент охватывает не только другие русские популяции (белорусов и украинцев), но и финно-угров, что говорит о единстве материнской «породы». Следует говорить не о финно-угорском субстрате в русском генофонде, а о дочернем характере финно-угров по отношению к русскому генофонду.
В пространстве главных компонент восточно-европейские популяции с несколько иным составом проанализированных этносов проявляется достаточно своеобразная закономерность. Если аварцы и мордва фактически представлены линейными закономерностями (то есть, для них может быть выделен единственный комплекс плавно меняющихся признаков), то белорусы явно представлены двумя подобными ветвями — одной, совпадающей с русской ветвью, и другой, ортогональной первой, — вне русского «генетического облака». Причем линейная ветвь мордвы выглядит продолжением белорусской ветви (плавным изменением той же тенденции трансформации генетических признаков). Мари образуют компактное облако с низкой изменчивостью. Башкиры сочетают одну доминирующую и одну второстепенную ветви, смешанные меж собой. Наконец, русское «генетическое облако» выглядит составленным из множества больших и малых линейных ветвей.
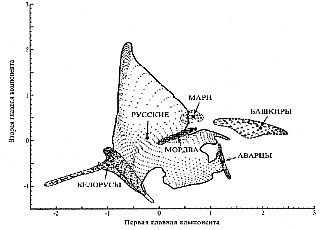
Изменчивость генофонда Восточной Европы в пространстве главных компонент (стрелками обозначены центры тяжести «этнических облаков»).
[Лимборская С.А. и др. Этногеномика и геногеография народов Восточной Европы. — М.: Наука, 2002.]
В предыдущих главах уже приводилось значение частот маркеров мтДНК для расовой диагностики.
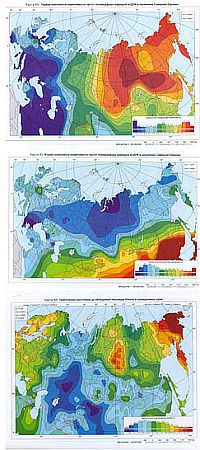
Пять главных компонент, рассчитанных для полиморфных маркеров мтДНК, охватывают 49 % совокупной изменчивости их частот. Первая главная компонента охватывает 18 % изменчивости, вторая — 11 %, третья — 7 %, четвертая — 7 %, пятая — 6 %. Визуально очевидна (и подтверждается корреляционным анализом) высокая степень сходства первых и вторых главных компонент мтДНК и аллельных генов. Также имеется достаточно ясное сходство между рельефом изменчивости пятой главной компоненты мтДНК и рельефом третьей главной компоненты изменчивости аллельных генов.
На рисунках мы ограничиваемся лишь двумя главными компонентами. Учитывая, что данные геногеографии по Зауралью серьезно искажены исключением русских, мы можем видеть обширные «низкочастотные территории вокруг Черного и Каспийского моря на первой главной компоненте, шлейф от Кавказа на второй главной компоненте и алтайско-южноуральский регион на третьей главной компоненте. Возможно, каждая из «ортогональных» групп изменчивости свидетельствует об уникальном векторе генетической эволюции: первая — о кавказоидной экспансии, третья — о южноуральской (арийской) вотчине, вторая (с меньшей уверенность) — о встречной экспансии с Кавказа и из Сибири и следах других разнородных миграций.
Первая и вторая главные компоненты изменчивости частот полиморфных маркеров мтДНК, генетические расстояния от усредненной (обобщенной) евразийской популяции
[Генофонд и геногеография народонаселения. Том 2. Геногеографический атлас населения России и сопредельных стран. СПб.: Наука, 2003.]
Западноевразийской (европейской) гаплогруппы I, U1, U2, U3, X у русских встречаются только в южной части русского ареала и отсутствуют в северных популяциях (Пинега, Каргополь). В целом средняя частота таких гаплогрупп у русских 1–2%. Напротив, частота западноевразийской гаплогруппы H у русских высока. Она падает с севера на юг от значений около 50 % в Архангельской области до 35–40 % у южных русских, в чем отражается общеевропейская тенденция. Восточноевразийские гаплогруппы у русских встречаются с низкой частотой и распределены в русском ареале повсеместно. Суммарная частота всех восточно-евразийских гаплогрупп в русских популяциях (2,2 %) низка, как и у народов Западной Европы (у французов в среднем 0,5 %). Для сравнения: такие гаплогруппы встречаются с частотой 85-100 % у монголов, китайцев, эскимосов.
В русских популяциях возможна тенденция некоторого нарастания восточноевразийских гаплогрупп к востоку (Унжа — 3,8 %). Загадочным является аномально высокая частота гаплогруппы D на северо-востоке (Пинега — 5,6 %, в целом по восточноевразийским гаплогруппам — 6,2 %). Причем с антропологической точки зрения это типично старорусское население: высокорослый северорусский тип с выраженной горизонтальной профилировкой лица, сильно выступающим носом и высоким процентом светло-голубых глаз и русых волос. Никакой связи с соседствующими коми в пинежской популяции не прослеживается, признаков родства с финно-угорскими народами — также.
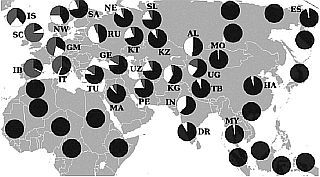
Многие гаплогруппы Y хромосомы, обнаруженные в коренных русских популяциях, редки и не удовлетворяют 1 % критерию полиморфизма. Среди гаплогрупп русского ареала имеют частоту выше 1 % только восемь. Они встречаются (порознь или вместе) более чем в 90 % случаев. По ним мы можем увидеть основные связи с иными ареалами.
Распределение частот гаплогрупп Y-хромосомы: R1a (белые сектора круговых диаграмм) и R1b (серые сектора). По гаплогруппе R1b выделяется ареал доарийского населения Европы, представленного более высокими частотами на западе Европы и следовыми количествами в Восточной Европе и в западной Азии. По гаплогруппе R1a прослеживаются последствия арийских миграций — в Индию, Персию, Европу, в незначительной доле — в Малую Азию, Аравию и на Кавказ. Обе гаплогруппы практически отсутствуют в Китае, Восточной Сибири, в Юго-Восточной Азии и в Африке.