Составим для этого прежде всего в общем виде энергетический баланс, характерный для растений, а затем такой же для животных. Такой баланс можно представить достаточно надежно, если не углубляться в существо сложнейших биологических процессов, а ограничиться входящими и выходящими потоками энергии.
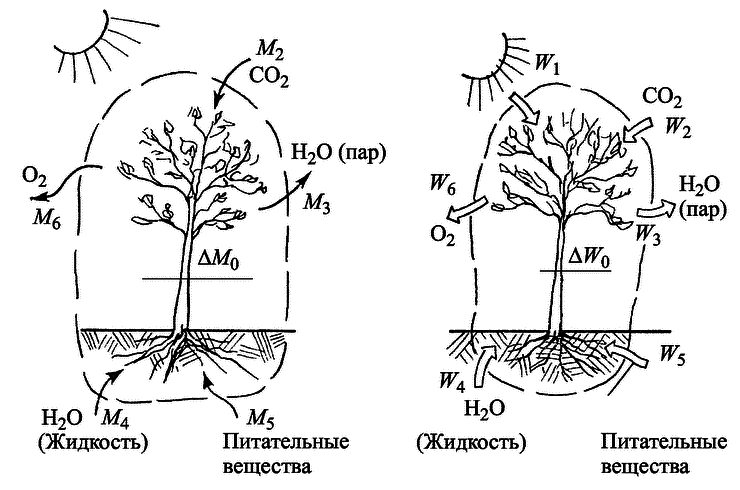
На рис. 4.3 представлена схема материального (потоки вещества) и энергетического балансов растения, основанных на законах сохранения массы и энергии. Чтобы составить такие балансы, окружим растение так называемой замкнутой контрольной поверхностью (штриховая линия), чтобы учесть все входящие и выходящие потоки. Если хотя бы один из них ускользнет от учета (или, наоборот, будет учтен тот, который через контрольную поверхность не проходит), баланс станет неверным. Тогда никаких мало-мальски стоящих выводов из него делать нельзя. Мы постараемся не допустить такой ошибки.
Рис. 4.3. Схемы материального (а) и энергетического (б) балансов растения
Материальный баланс будет иметь вид
М2 + М4 + М5 — (М3 + М6) = ΔМ0.
Это уравнение показывает: все, что получает растение (М2 + М4 + М5) за определенный отрезок времени, за вычетом того, что оно отдает (M3 + M6) идет на приращение ΔМ0 его массы, связанное с ростом. Аналогичное уравнение получится и для энергии:
W1 + W2 + W4 + W5 — (W3 + W6) = ΔW0.
Здесь ΔW0 — прирост внутренней энергии растения, определяемый увеличением его массы при росте.
Чтобы установить, нарушает эта система второй закон термодинамики или нет, нужно проверить, что происходит с энтропией в процессе жизнедеятельности растения: увеличивается она или уменьшается?
Очевидно, что живая ткань растения более высоко структурно организована, чем поступающие из воздуха питательные вещества. Поэтому при образовании такой ткани (с массой ΔM0) ее энтропия будет несомненно меньше, чем суммарная энтропия исходных веществ (СО2, H2O) и питательных веществ почвы). В этом смысле образование и накопление живой ткани растения и поддержание ее существования будет, несомненно, антиэнтропийным процессом. Но никак нельзя забывать, что одновременно неизбежно меняется энтропия потоков вещества и энергии, проходящих через контрольную поверхность. Здесь получается обратная картина (рис. 4.3): суммарная энтропия выходящих потоков (3 и 6) неизбежно оказывается много большей, чем входящих (7, 2, 4 и 5). Это объясняется тем, что энтропия поглощаемого солнечного излучения[60] сравнительно невелика, так же как и поступающих из почвы минеральных солей; энтропии газов — кислорода и СО2 — близки по значениям. Зато энтропия водяного пара, отдаваемого листьями, относительно велика (примерно в 3 раза больше, чем у воды). В результате энтропия потоков, проходящих через контрольную поверхность, возрастает намного больше, чем снижается энтропия веществ, превращающихся в органическую ткань.
Если первую величину — прирост энтропии — обозначить через ΔS', а вторую (уменьшение энтропии) — через ΔSʺ, то оказывается, что всегда ΔS' >> ΔSʺ.
Следовательно, в целом энтропия неизбежно возрастает на величину
ΔS = ΔS' — ΔSʺ >> 0.
Другими словами, растения только потому могут расти антиэнтропийно, что «сбрасывают» избыток энтропии в окружающую среду; при этом прирост энтропии в ней намного больше, чем снижение ее в самом растении. Поэтому все рассуждения об «антиэнтропийной природе растительной жизни», о «нарушении второго закона термодинамики» совершенно неверны. Они основаны на неполном учете всех величин, определяющих общее изменение энтропии, подмене точного анализа и расчета общими рассуждениями.
Если взять другую часть органического мира — животных, то здесь наблюдается та же картина. Животные, питаясь растениями (или другими животными), а также поглощая воду и кислород воздуха, выделяют СО2, теплоту и продукты, получаемые в результате переваривания пищи. Энтропия всего того, что выделяется, намного больше энтропии того, что поглощается. В результате уменьшение энтропии, происходящее как при образовании новых органических тканей и отмирании старых, оказывается намного меньше, чем общий прирост энтропии. Животные тоже «сбрасывают» излишнюю энтропию в окружающую среду, развивая или поддерживая таким путем свою внутреннюю высокоорганизованную малоэнтропийную структуру. В целом энтропия опять неизбежно растет. Очень наглядно проиллюстрировал это положение Э. Шредингер, о котором мы уже упоминали. Он писал так: «Энтропия кошки уменьшается за счет того, что возрастает энтропия системы «кошка + мышь»; т. е. то, что получается из мыши после того, как кошка ее съест и переварит, имеет значительно большую энтропию, чем мышь».
Таким образом и с другой, биологической стороны опровергнуть второй закон тоже не удается. Остается еще одна, последняя возможность — создать техническое устройство (или найти готовое), которое действовало бы вопреки второму закону термодинамики. Лучше всего, конечно, было бы, если бы такая система была двигателем и производила работу; но это в конце концов необязательно. Для доказательства достаточно только указать любую систему такого рода, поскольку возможность ее, существования однозначно определяет и возможность создания действующего ppm-2. В последнем параграфе этой главы мы рассмотрим одно такое устройство — тепловой насос, принцип действия которого уже обсуждался на стр. 125. По мнению многих сторонников «энергетической инверсии», он своей работой наилучшим образом опровергает второй закон термодинамики.
Прежде чем приступить к разбору теплового насоса, полезно проделать небольшую работу по ознакомлению с одним термодинамическим методом, который позволяет просто и наглядно определять, может ли работать любое предложенное устройство с точки зрения второго закона термодинамики, и если может, то какова его термодинамическая эффективность. Это не только очень удобно для анализа теплового насоса, но и позволит дополнительно рассмотреть роль второго начала термодинамики в биологии.
4.3. Эксергетический баланс и КПД
Энтропия — основная величина, определяющая возможность (или невозможность) протекания процессов в любых системах преобразования вещества и энергии с позиций второго закона термодинамики. Суммарная энтропия неизменна или растет — процесс возможен; уменьшается — невозможен. В рассмотренных выше случаях мы успешно пользовались именно этим фундаментальным свойством энтропии для того, чтобы определить, что может быть в энергетических превращениях и чего быть не может. Однако не только этим свойством определяются возможности практического использования энтропии. Она может помочь в решении другой, не менее важной задачи — определить качество энергетических превращений (а следовательно, и любых устройств, в которых они производятся).
Когда говорят об энергосберегающей технологии, об уменьшении потерь энергии, то речь по существу идет не об энергии в количественном смысле. Нужно четко понимать, что в количественном отношении энергию сберегать не нужно, об этом автоматически заботится первый закон термодинамики — закон сохранения энергии. Любое техническое устройство (да и вообще все на свете, что живет и движется) действует всегда так, что энергия сохраняется: сколько ее входит, столько неизбежно и выходит; она никогда не теряется. Поэтому сбережение энергии — это по существу сбережение ее качества. Именно об этой качественной стороне энергии писал Энгельс в «Диалектике природы». Любая технология и технические устройства, в которых она осуществляется, тем совершеннее, чем меньше будет возрастать энтропия в результате их функционирования, т. е. чем меньше будет «портиться» энергия.