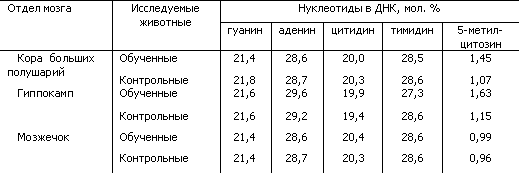
Таблица 5. Содержание нуклеотидов и метиловых компонентов в ДНК из разных отделов мозга у обученных и контрольных крыс (по Н.А. Тушмаловой)
Как уже было упомянуто, основные структуры нервной клетки, реализующие ее функции, состоят главным образом из белковых соединений. Поэтому многие исследователи ведут изучение белков как возможного субстрата долговременной памяти. Усиленный синтез белка при обучении дает основания для суждений о его роли в явлениях памяти. Чтобы выяснить, какие именно белки усиленно синтезируются для обеспечения процессов фиксации памятного следа, предпринималось изучение их раздельных фракций.
Исследование белков мозга голубей выявило, что при их обучении происходит возрастание фракции мембранных гликопротеидов, обозначенных 10B и 11A. Эти белки благодаря своим углеводным компонентам могут регулировать активность возбудимых мембран и таким образом участвовать в процессах памяти. При обучении крыс применять непредпочитаемую лапу для добывания пищи последующее дробное фракционирование мозговых белков показало особое положение встречающихся только в нервной ткани белков S-100 и 14-3-2. Конформационные изменения S-100 при связывании с ионами Ca влияют на проницаемость мембран, в частности синаптических, определяя их функциональное состояние.
Значение белков в механизмах формирования долгосрочной памяти продемонстрировано в опытах, где угнетение синтеза белка нарушало переход краткосрочной памяти в долгосрочную. Так, введение ингибитора белкового синтеза пуромицина золотым рыбкам сразу после обучения стирало его следы, но введение пуромицина через 1 ч не вызывало такого действия. Эти опыты были повторены и на других животных, показывая, что синтез белка необходим для консолидации памяти, а когда памятный след закреплен, белок уже не столь нужен.
Какие же именно белки разрушаются под действием ингибитора, препятствующего формированию долговременной памяти? Для ответа на этот вопрос исследовали разные фракции мембранных белков синаптического аппарата после нарушения долговременной памяти пуромицином. Показано, что этот ингибитор наиболее резко тормозит синтез кислых белков, к которым относится специфический для тканей мозга белок S-100. Оказалось также, что белки наиболее чувствительной к ингибитору фракции по своему аминокислотному составу принадлежат к группе внутренних белков мембраны, которые связаны с ее функциями как холинорецепторы. Это соответствует мнению, что носителями долгосрочной памяти являются холинэргические синапсы. Однако имеются указания на участие в механизмах долгосрочной памяти и адренэргической системы. Отмечено, что положительное подкрепление условных рефлексов, например пищевых, связано с функционированием преимущественно серотонинэргических, а отрицательное при оборонительных рефлексах — норадренэргических структур мозга. О возможных нейрохимических компонентах подкрепления существуют разные мнения: сюда относят и норадренэргическую систему, и дофаминэргическую. За последнее время накоплено большое количество фактов, показывающих, как участвуют нейрохимические механизмы в процессах обучения и памяти.
Другой подход к выяснению роли белков в долговременной памяти состоял в прослеживании включения в них меченых аминокислот и изменений этих белков при обучении. Так, внедрение меченного тритием лейцина в белки мозга крыс после обучения возрастало и сопровождалось увеличением числа рибосом, связанных с мембранами. При переучивании крыс способу добывания пищи на стадии бурной двигательной активности происходило даже снижение включения меченого лейцина в белки мозга. По мере усвоения нового навыка включение меченого лейцина в белки, особенно специфические, возрастает. После выработки нового навыка п<...пропуск части слова...>ния у золотых рыбок отмечен синтез трех новых фракций белков, обнаруженных с помощью меченого валина. У крыс, обученных дифференцированию зрительных сигналов для решения лабиринтной задачи, наблюдали ускоренное включение меченого тирозина в мембранные белки. Выработка условных рефлексов у голубей сопровождалась усилением синтеза гликопротеинов по показателям включения меченого валина.
В последние годы все более широко применяют методы иммунохимической идентификации специфичных для мозга белков, в частности S-100, 14-3-2; гликопротеинов для определения их роли в фазировании свойств возбудимых мембран и синаптических процессов. Таким образом, накапливаются веские доказательства непосредственного участия белков, особенно кислых мембранных и гликопротеинов в переходе краткосрочной памяти в долгосрочную.
Сравнительно недавно было выяснено, что к возможным носителям долгосрочной памяти относится ряд пептидов — веществ, как и белки, состоящих из аминокислот, но более простых по своему строению. Такие пептиды ускоряют формирование условных рефлексов. Их действие на другие организмы воспроизводит выработанный навык, что рассматривается как явление «переноса памяти» (см. ниже).
К этим пептидам относятся главным образом такие гормоны гипофиза, как адренокортикотропин, меланоцитостимулирующий гормон, вазопрессин, окситоцин и некоторые рилизинг-гормоны; а также пептиды, специфически связанные с обучением какому-либо одному определенному навыку. Связь пептидов с памятью, по-видимому, определяется их участием в синаптической передаче. Предполагают даже существование особых «пептидэргических синусов», которые могут изменять свои свойства проведения.
Кроме того, считают, что факторы пептидной природы могут участвовать в образовании временных связей как химический механизм воздействия подкрепляющих систем на генетический аппарат нервных клеток.
Одно из замечательных свойств памяти состоит в специфической избирательности и локальности образуемых временных связей. В хорошо выработанном условном рефлексе каждый сигнал вызывает только свою строго определенную реакцию. В этом, а также в длительности хранения, свойствах угасания и восстановления, в чувствительности к ядам, тормозящим синтез белков и РНК, «память» нервной системы очень напоминает «память» системы иммунитета. Предположение об участии иммунных реакций в формировании долгосрочной памяти получило новые подтверждения и привело к разработке гипотезы об иммунохимическом механизме формирования долговременной памяти (И.П. Ашмарин, 1973).
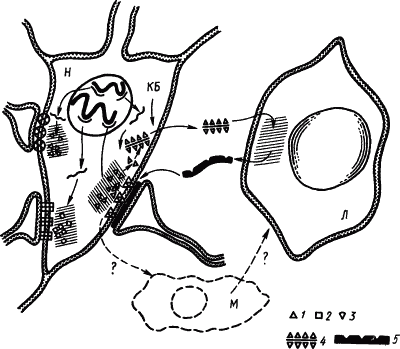
Согласно этой гипотезе в результате метаболизма синаптических мембран на их поверхности образуются вещества, которые, накапливаясь в избытке, при реверберации импульсов в стадии краткосрочной памяти играют роль антигена для антител, вырабатываемых в глиальных клетках. Связывание антитела с антигеном происходит при участии активаторов образования медиаторов или ингибитора ферментов, расщепляющих эти активаторы (рис. 26). В пользу этой гипотезы свидетельствуют эксперименты, показавшие, что антитела к синаптосомам повышали чувствительность последних к медиаторам и что стимуляторы иммуногенеза улучшали долгосрочную память.
Рис. 26. Иммунологические механизмы, повышающие проводимость определенных синапсов (по И.П. Ашмарину):
КБ — катионные белки, открывающие путь комплексам фрагментов антигена синапса с РНК через оболочку клетки; Л — клетки глии, функционально подобные лимфоцитам, М — клетки глии, функционально подобные макрофагам; 1, 2, 3 — фрагменты синаптических антигенов (зоны их синтеза заштрихованы), 4 — комплекс фрагмента антигена с РНК, 5 — антитела для синапсов, тонкими линиями изображена мРНК, толстыми — ДНК хромосом
Участие в иммунохимических механизмах памяти специфичного для мозга белка S-100 показано в опытах эпидурального введения крысам сыворотки с антителами к этому белку, в результате чего затруднялась выработка навыка поведения в лабиринте. Чтобы выделить в «чистом виде» эффект действия антител на белки, специфические для процессов образования временных связей, изучали навык поведения крыс. При этом вводили антитела, которые вырабатывались на гомогенаты мозга необученных и обученных крыс, а также крыс, получавших препараты, облегчающие или затрудняющие обучение.