Эффект действия антител резко проявлялся уже в 1-й день обучения в виде угнетения условных реакций животных, получавших антитела к веществам-участникам формирования долговременной памяти под влиянием препаратов, облегчающих обучение. Видимо, такой эффект объясняется связыванием и дезактивацией этих веществ.
Больше половины мозга занимают глиальные клетки, которых примерно в 10 раз больше, чем нервных; каждый нейрон со всех сторон окружен глией. Естественным было предположение, что глия участвует в осуществлении функций мозга, в том числе высшей нервной деятельности. Замедленность реакций клеток глии и сосредоточение в них специфичных для мозга белков S-100, некоторых гликопротеинов и биологически активных соединений создают условия для реализации постепенно стабилизирующихся процессов, к каким относится память. Поэтому появились основания предполагать, что клетки глии во взаимодействии с нейронами принимают деятельное участие в формировании долгосрочной памяти.
Для подтверждения этого предположения проводили эксперименты, показавшие, что инъекции экстрактов глии крысам ускоряли выработку условных рефлексов. В пользу такого предположения свидетельствует и прогрессивное увеличение глиальной ткани относительно нервной в ряду от низших животных к высшим. Медленная электрическая активность глии может быть источником колебаний постоянного потенциала мозга и электротонических потенциалов, создающих очаги повышенной возбудимости, которые считают основой образования временных связей.
О возможном механизме участия глии в фиксации памятного следа существуют различные мнения. Так, предполагали, что выходящие при возбуждении нейрона ионы K захватываются клетками глии и передаются следующему нейрону, вызывая стойкое изменение уровня его поляризации, а следовательно, облегчение его возбуждения. Другое объяснение состоит в том, что выработка условного рефлекса вызывает у прилегающих к нейрону глиальных клеток усиленное образование миелина, который окутывает его тончайшие терминальные волокна и уменьшает фактор риска проведения по ним импульсов, в результате чего «потенциальные» синапсы становятся действующими (А.И. Ройтбак, 1968). Схема происходящих при этом процессов представлена на рис. 27. Эта гипотеза получила экспериментальное подтверждение в стимуляции миелинообразования глиальными клетками с помощью ионов K, которые выходят из возбужденного нейрона в межклеточную среду.
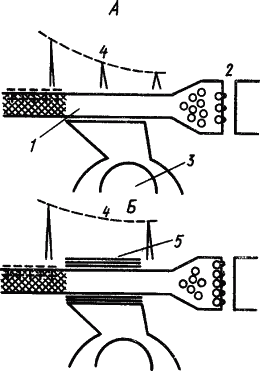
Рис. 27. Гипотеза о роли глии в образовании временной связи условного рефлекса. А — синапс не проводит, так как потенциал возбуждения в немиелинизированной пресинаптической терминали ослабляется и медиатор выделяется в количестве, недостаточном для срабатывания синапса; Б — глиальная клетка, возбуждаясь при безусловном подкреплении рефлекса, замыкает пресинаптический участок терминали в миелиновую оболочку и потенциал возбуждения сохраняет величину, необходимую для срабатывания синапса (по А.И. Ройтбаку):
1 — аксон, 2 — синапс, 3 — глиальная клетка, 4 — изменение потенциала действия, 5 — миелиновая оболочка
Для того чтобы определить, какова в долговременной памяти роль глии и нейронов, испытывали действие на условные рефлексы крыс антител, полученных при иммунизации гомогенатом или глии, или нейронов. Оказалось, что антинейрональная сыворотка ухудшала главным образом краткосрочную память, а антиглиальная — долгосрочную. Эти результаты рассматривают как указание на то, что краткосрочная память обусловлена реверберацией импульсов по цепям нейронов, а долгосрочная — реализуется пластическими процессами с участием глии.
Уже давно высказали мнение, что нервные клетки выполняют свои функции в виде объединений. Таковы динамические констелляции (А.А. Ухтомский, 1945) и образующиеся при обучении функциональные ансамбли (Д. Хебб, 1949). Экспериментальные исследования показали реальность существования таких объединений, названных элементарными нейронными ансамблями (А.Б. Коган, 1964, 1979). Нейронные ансамбли обладают свойствами, отличными от свойств отдельных нейронов.
Качественное своеобразие системной деятельности нейронов обосновывается морфологическими особенностями их связей (Я. Сентаготаи, 1968), эффектами объединения; существует представление о том, что при обучении временные связи формируются путем объединения нейронов различных структур мозга в комплексы, называемые микросистемами (М.М. Хананашвили, 1972). Наименьшие микрообъединения нейронов коры, строящие свою деятельность на основе предыдущего опыта, обозначают как нейроценозы. Локальная аппликация ацетилхолина в качестве подкрепления выявляла группы нейронов, проявляющие синхронные пластические перестройки. Эти нейроны определяли как микропулы.
Существует концепция, по которой каждый акт высшей нервной деятельности связан с активацией определенной совокупности нейронов, образующей сложную мозаику. Такой подход к проблеме памяти лишен затруднений, возникающих перед гипотезами ее «молекулярного кодирования», и подкрепляет концепцию «распределенной памяти», согласно которой механизмы фиксации памятного следа не сводятся к изменениям свойств отдельных клеток, а состоят в избирательном формировании определенных межнейронных отношений в их популяциях. Об этом свидетельствуют и результаты опытов с так называемым обучением по обратным связям, когда подкреплялась импульсация одного из трех одновременно регистрируемых нейронов слуховой коры кошки. Корреляционный анализ показал, что нейрон, импульсация которого служила сигналом, устанавливал определенную систему связи с остальными нейронами и такие межнейронные отношения могли сохраняться до 20 мин. Представление о совместной деятельности нейронных объединений легло в основу теории статистической конфигурации (Е. Джон, 1973), согласно которой носителем памятного следа является статистически формируемый набор активных нейронов, образующих фигуру определенных очертаний.
Ha основании большого экспериментального материала о взаимосвязи ритмических свойств нейронов, образующих цепочки возбуждения разной лабильности и определяющих функциональное состояние центральных нервных структур, было высказано мнение, что новая временная связь запечатлевается в нервной системе в виде специфического пространственно-временного распределения активности нейронов, перестроивших свою лабильность (М.Н. Ливанов, 1975). В пользу этого предположения свидетельствуют результаты исследования конфигурации мозаики нейронной активности, показавшие, что условный рефлекс проявляется лишь тогда, когда рисунок мозаики, вызванной сигналом, соответствует таковому памятных следов и подходит к нему, как «ключ к замку». Возникновение разных рисунков нейронной активности с участием одних и тех же нейронов означает перераспределение свойств проводимости их синапсов, которое может осуществляться рассмотренными выше механизмами.
Такое представление близко к взглядам на нейронную структуру условного рефлекса, как на массовое объединение нейронов, в котором воспроизводится вся ситуация в целом (А. Аттли, Т. Блумфилд, Е. Джон, 1973). Обобщение большого материала многолетних нейрохимических исследований памяти также привело к заключению о ее распределенном характере в ансамблях нейронов. При этом роль биохимических процессов, создающих сложные «рисунки» распределения химизма нервных клеток, заключается в организации «узнавания» клетками друг друга и «сборки» их в функциональные ансамбли. Существует предположение о химической природе сигнала, управляющего проходимостью синапсов нейронов. Это выражается перестройкой классов доминирующих межимпульсных интервалов, в активности нейронов.
Получены также факты, указывающие на то, что механизм образования условного рефлекса не ограничивается перестройками центральных процессов, а включает в себя и периферические гормональные компоненты. Особое значение при этом имеет система гипофиз — надпочечники и образование энкефалиноподобных пептидов. Эксперименты с действием мет- и лей-энкефалина на условные рефлексы избегания у крыс привели к предположению об их возможной роли в обучении и памяти.