Если относить к внутреннему все виды торможения, которые вырабатываются, то к нему принадлежит и торможение, возникающее при многократном повторении любого афферентного раздражения, которое приводит к угашению ориентировочной реакции на него. Такие явления «привыкания», или «габитуации», в сущности представляют собой случаи угашения натурального условного рефлекса. За последнее время они детально изучены на нейронном и молекулярном уровнях на гигантских нейронах беспозвоночных. При этом найдено, что угашение реакции втягивания жабер у аплизии сопровождается снижением моносинаптических ВПСП мотонейрона в результате уменьшения выброса медиатора из пресинаптических терминалей в синаптическую щель. Исследование квантового состава выделяющегося в синапс медиатора на примерах привыкания нервных клеток виноградной улитки и нейронов коры и гиппокампа кролика показало, что во всех случаях имело место уменьшение числа, а в некоторых случаях и объема квантов.
Несмотря на обилие работ по изучению внутреннего торможения, остается еще много неясного в его механизмах.
Глава 5
ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Вся сложная и разнообразная деятельность высших отделов нервной системы построена на работе двух основных нервных процессов — возбуждения и торможения. Протекая в подвижных пространственных и временных отношениях друг с другом, эти процессы то разливаются (иррадиируют), то сосредоточиваются (концентрируются), то возбуждение порождает торможение (отрицательная индукция), то торможение дает начало возбуждению (положительная индукция). Непрерывное взаимодействие движущихся и вызывающих друг друга возбудительных и тормозных процессов создает в высших отделах мозга чрезвычайно тонкую мозаику, колеблющийся узор из переплетения возбужденных и заторможенных нейронов. Такие мозаики лежат в основе как различных актов поведения, так и их торможения в явлениях сна.
Иррадиация и концентрация нервных процессов
Возбуждение или торможение, возникшие в какой-либо клетке или группе клеток мозга, не остаются неподвижными, а всегда склонны к распространению. Распространение нервного процесса из очага его возникновения на окружающие нервные клетки называется иррадиацией.
Иррадиация торможения. Иррадиацию условного торможения удобно наблюдать в кожном анализаторе. Широко развернутая кожная поверхность тела является как бы увеличивающим зеркалом, в котором можно ясно видеть, как по последовательно расположенным проекционным полям будет иррадиировать тормозное состояние, например дифференцировочное торможение.
Иррадиацию дифференцировочного торможения обнаружили в следующем опыте.
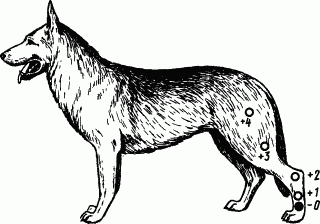
Вдоль задней ноги собаки от стопы до бедра наклеивали пять «касалок» — приборов для механического раздражения кожи (рис. 35). Четыре верхние касалки использовали как сигналы условных пищевых слюноотделительных рефлексов. Эти рефлексы укрепляли, пока их слюноотделительный эффект не стал приблизительно одинаковым. Нижняя касалка служила дифференцировочным раздражителем и применялась без подкрепления едой, пока не перестала вызывать даже малейшего слюноотделения. Если теперь, вслед за применением дифференцировочной касалки, попробовать положительные раздражители, то оказывается, что слюногонное действие последних претерпевает закономерные изменения.
Рис. 35. Опыт с иррадиацией дифференцировочного торможения по корковым клеткам кожного анализатора:
— дифференцировочный раздражитель, 1, 2, 3, 4 — положительные условные раздражители (прикладываются к пунктам кожи ноги на расстоянии от дифференцировочного раздражителя соответственно на 3, 9, 15 и 22 см)
В классических опытах Н.Н. Красногорского и Б.А. Когана условный рефлекс на каждую из положительных касалок составлял 5 капель слюны за 30 с. Но каждый раз, когда дифференцировочная касалка создавала очаг торможения, начинали изменяться и соседние положительные рефлексы. Следовательно, торможение выходит за пределы своего очага и захватывает соседние клетки анализатора, в данном случае те, на которые проецируются кожные пункты положительных касалок. Далее видно, что в одинаковых условиях минутного интервала после троекратного действия дифференцировочной касалки условные рефлексы с положительных касалок изменяются по-разному, в зависимости от расположения последних относительно дифференцировочной. Так, рефлекс с ближайшего пункта (касалка 1) оказался полностью заторможенным. Рефлекс с пункта, расположенного несколько дальше (касалка № 2), был только уменьшен. Рефлексы с остальных пунктов, расположенных еще дальше, не только не испытали торможения, но даже усилились. Следовательно, иррадиирующее торможение оказывает тем более сильное воздействие на клетки анализатора, чем ближе они находятся к тормозному очагу. По своей природе иррадиация торможения — это развитие ТПСП на все большей территории коры, но не истинное движение процесса.
Концентрация торможения. После широкой иррадиации наступает сосредоточение, концентрация торможения в месте своего возникновения. Этот процесс тоже удобно проследить на примере дифференцировочного торможения в кожном анализаторе. Опыты проводили так же, как и при наблюдении иррадиации, но положительные рефлексы с каждого пункта кожи испытывали в различные сроки после окончания действия тормозного раздражителя. При помощи такого приема можно увидеть, как сначала далеко распространившееся тормозное состояние начинает сосредоточиваться, возвращаясь к исходному пункту.
При своем концентрировании торможение проходит в обратной последовательности все те пункты проекционных полей анализаторов, которые оно захватывало в своем поступательном движении.
Что собой представляет процесс концентрации торможения? Здесь не исключены две возможности. Первая заключается в том, что распространившееся торможение рассеивается, затухает на периферии и занимаемая им территория постепенно уменьшается. Вторая возможность — это подъем обратной волны торможения к тому месту, откуда оно распространилось. Последнее более вероятно, как, например, упрочение дифференцировки сопровождается усилением тормозного процесса. Следовательно, концентрация торможения связана не с рассеиванием и ослаблением, а с его сосредоточением и усилением.
Скорость иррадиации и концентрации торможения в коре головного мозга собаки. Скорость иррадиирования тормозного состояния удалось измерить в кожном анализаторе. Для этого определяли время развития вторичного торможения условного рефлекса на раздражение пункта кожи, находящегося в известном удалении от первично угашаемого. Опыты проводили следующим образом. В разных местах тела собаки приклеивали к коже касалки. На все касалки вырабатывали условные пищевые слюнные рефлексы и укрепляли их до одинаковой величины. Затем наносили повторные раздражения какой-либо одной касалкой без подкреплений до тех пор, пока не достигали исчезновения слюноотделения. Как только эта касалка проявляла свое тормозное действие, испытывали состояние рефлекса с другого пункта. На основании ряда таких испытаний в разные сроки после вызова угасательного торможения и при разных расстояниях испытуемых пунктов от его очага определяли время иррадиирования (Б.А. Коган).
В этом опыте вторично затормаживаемые пункты испытывали медленно после раздражения, давшего нулевой результат, т.е. без всякого промежутка времени между ними. В этот момент, как видно, торможение из пункта, соответствующего правой стороне груди еще не успело распространиться дальше ближайших участков. До области, например, правой предплюсны оно еще не дошло. Однако постепенно увеличивая промежутки времени от тормозного раздражения до момента испытания рефлекса, можно уловить момент, когда торможение дойдет до пункта, соответствующего правой предплюсне. Оказалось, что процесс иррадиирования торможения по нервным клеткам коры протекает очень медленно. Для прохождения области одного только кожного анализатора торможению требуются минуты.