Рис. 33. Реакция нейронов коры мозга обезьяны, отвечающих возбуждением (+) или торможением (−) на действие положительного сигнала с электрокожным подкреплением (заштрихованные столбики) и дифференцировочным без подкрепления (белые светлые столбики) (по Г. Джасперу и др.):
I — моторная зона, II — зрительная, III — теменная, IV — лобная
Развитие внутреннего торможения, например, при угашении условных рефлексов проявлялось в том, что у нейронов, реагирующих возбуждением, наблюдали снижение частоты условного разряда и даже замедление фоновой импульсации. У нейронов, реагирующих торможением, отмечали повышение частоты разряда на отрицательный сигнал. Наглядная иллюстрация такой динамики нейронных реакций представлена на рис. 34.
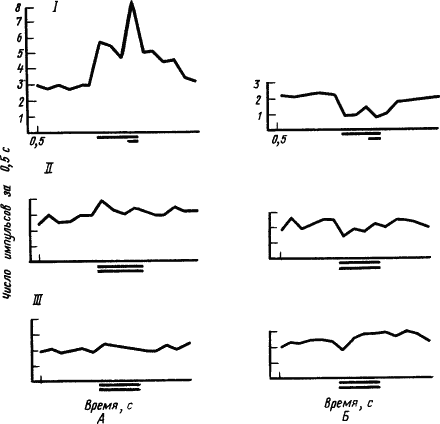
Рис. 34. Динамика реакции I, II, III возбуждения (А) и торможения (Б) нейронов двигательной коры кролика при выработке дифференцировочного торможения (по Е.Л. Полонской):
длинной линией под абсциссой обозначено время действия условного раздражителя, короткой линией под ней — <...> безусловного, двумя длинными линиями — действие условного раздражителя без безусловного
Одна из особенностей тормозных реакций нейронов состоит в том, что они угашаются быстрее, чем возбудительные. Так, в опытах с угашением условных рефлексов показано, что в то время как условные реакции возбуждающихся нейронов угашались постепенно и не полностью, реакции тормозящихся нейронов быстро исчезали. При этом наиболее устойчивыми оказывались те фазы импульсного ответа нейрона, которые соответствовали времени бывшего подкрепления. Отмечены существенные различия реакций нейронов разных отделов мозга на развитие внутреннего торможения. Например, нейроны ретикулярной формации среднего мозга крысы при неподкреплении сигнала пищевого рефлекса очень быстро перестают него реагировать, а нейроны гиппокампа еще продолжают отвечать, лишь уменьшив частоту импульсов. Указывают, что импульсные ответы нейронов гиппокампа обладают более высокой точностью дифференцирования по сравнению с другими подкорковыми структурами. Особенности динамики нейронных реакций при выработке условного торможения описаны при исследовании слуховой и моторной коры, а также внутреннего коленчатого тела у кроликов.
В опытах на зрительной коре морской свинки было показано, что тормозные влияния осуществляются от периферических частей микроансамблей нейронов к их центральным частям.
По поводу нейронов, тормозящихся даже при осуществлении условных рефлексов, было высказано предположение, что они представляют особую форму внутреннего торможения, которая сопутствует каждому акту условно-рефлекторной деятельности, и обеспечивают его приспособительную точность. Однако такое определение роли тормозящихся нейронов встретило возражение, согласно которому в описываемой ситуации речь идет не о внутреннем, а о координационном торможении, обеспечивающем точность выполнения условного рефлекса.
Таким образом, условное торможение проявляется в специфической организации возбужденных и заторможенных нейронов, участвующих в создании временной связи, и различных их взаимоотношениях в разных структурах коры и подкорковых образованиях мозга.
Нейрофизиологический механизм внутреннего торможения. О нейрофизиологии внутреннего торможения высказывались разные предположения. Так, И.П. Павлов (1927), не решая окончательно вопрос о его природе, считал возможным рассматривать развитие внутреннего торможения при неподкреплении как результат перевозбуждения клеток коркового представительства сигнала, которые при подкреплении предохранялись от этого затормаживанием. Исследование свойств угасательного торможения привело к сходному заключению о том, что оно является фазой перевозбуждения возбудительного очага. Другое предположение исходило из того, что при неподкреплении ослабляются прямые временные связи от зоны корковой проекции условного раздражителя к зоне безусловного и усиливаются обратные, по которым активируются апикальные дендриты выходных пирамидных нейронов в зоне представительства безусловного рефлекса. Такое локальное возбуждение дендритов создает электротоническую блокаду синапсов, и рефлекс прекращается (И.С. Беритов, 1956). Другим источником локального возбуждения апикальных дендритов пирамид могут быть неспецифические структуры таламуса (А.И. Ройтбак, 1958).
Исходя из того, что рабочее состояние высших отделов мозга обусловливается тоническим восходящим активирующим влиянием ретикулярной формации среднего мозга, условное торможение рассматривали как результат снижения этого влияния. Дальнейшие исследования показали, что для развития внутреннего торможения. Важное значение имеет изменение тонуса ретикулярной формации (Б.И. Котляр, 1977). Однако это не единственная причина внутреннего торможения.
По другому предположению, ведущую роль в угнетении условных рефлексов играет антагонист мезэнцефалической системы — восходящая «синхронизирующая» таламокортикальная система (Г. Мегун, 1956). Считают также, что поскольку автостабилизация корковых процессов поддерживается «превентивным» торможением (П.В. Симонов, 1965), его углубление при действии неподкрепляемых раздражений образует внутреннее торможение.
На основании сходства ряда электрофизиологических показателей реализации положительных и отрицательных условных рефлексов, особенно при тонких дифференцировках, условное торможение рассматривают как возбуждение, ослабленное отсутствием обратной афферентации от исполнительных органов ввиду отсутствия подкрепления (Ю.Г. Кратин, 1967). Неподкрепление условного рефлекса само по себе становится сигналом «неблагополучия» и создает «трудное состояние», возникновение которого по механизму внешнего торможения задерживает рефлекс (П.К. Анохин, 1968). Различия порогов реакций, вызываемых безусловным и условным раздражителем, объясняют частичным торможением центральных структур собственного рефлекса на раздражитель, который сделали условным, а отмена подкрепления нарушает этот механизм, вызывает перевозбуждение и охранительное торможение в элементах временной связи (Э.А. Асратян, 1970).
Существует мнение, что внутреннее торможение есть не что иное, как образование нового рефлекса, в котором подкреплением является отсутствие безусловного раздражителя (Ю. Конорски, 1970). Условное торможение по многим своим проявлениям и электрофизиологическим показателям обнаруживает свойства доминанты, которая становится очагом нераспространяющихся возбуждений (В.С. Русинов, 1969). Если образование условного рефлекса происходит путем «прокладывания связей» между сигналом и доминантным очагом безусловного рефлекса, то при отсутствии подкрепления предполагают накопление в представительстве сигнального стимула нераспространяющегося возбуждения, образующего своеобразный локальный доминантный очаг. Поскольку образование пути условного возбуждения определяется пространственной синхронизацией и когерентностью ритмов активности (М.Н. Ливанов, 1972), их рассогласование препятствует распространению возбуждения по путям временных связей. Этому соответствуют данные о том, что при выработке дифференцировки двигательного рефлекса на звук коэффициенты корреляции импульсных потоков нейронов слуховой и ассоциативной коры кошки ранее положительные становятся отрицательными.
На основании результатов опытов, в которых изучали влияние симпатической части нервной системы на высшую нервную деятельность, был сделан вывод о ее участии в механизме формирования внутреннего торможения. Было высказано предположение, что выработка дифференцировочного торможения представляет собой образование нового рефлекса с подкреплением из памяти. Исследование процессов внутреннего торможения на нейронном и системном уровнях привело к заключению о существенной роли в его механизме явлений возвратного торможения и развития гиперполяризационных процессов в коре. Недавно представлены факты, свидетельствующие о том, что при изучении механизма внутреннего торможения следует учитывать изменения тонуса активности нейронов коры и подкорковых структур головного мозга.