Кроме холинэргических имеются и иные синапсы, где медиаторами служат другие вещества. С учетом всех этих сведений была составлена обобщенная схема стойкого облегчения медиаторных процессов в синапсе. На рис. 24, А показано исходное состояние синапсов. Если возбуждение пресинаптического нейрона (рис. 24, Б) вызывает усиленное выделение медиатора, а возбуждение постсинаптического — разрушающего медиатор фермента (рис. 24, В), то при сочетаниях условного и безусловного раздражителей происходит конвергенция импульсных потоков на синапсах (рис. 24, Г), которая обусловливает оптимальный баланс активности медиатора и разрушающего его фермента на стабильном высоком уровне, а также активацию сети белковых тяжей, уменьшающих просвет синаптической щели (рис. 24, Д). Показано, что повышение эффективности синапсов сопровождается конформационными изменениями мембранных белков, специфичными для синаптических мембран.
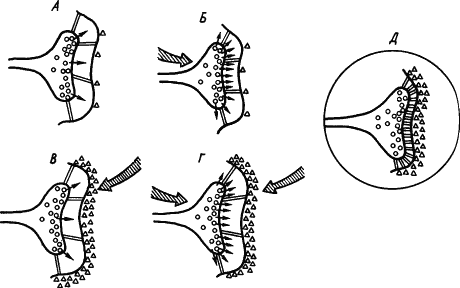
Рис. 24. Развитие процессов стойкого облегчения проводимости сигнала (объяснение см. в тексте) (по В.В. Дергачеву):
кружками обозначены везикулы медиатора, треугольниками — фермент, расцепляющий медиатор
Долгосрочная память как стойкое изменение химизма нервных клеток. Проведение сигналов от клетки к клетке, вызывающее активацию нейронных популяций, происходит в синапсах, однако функциональное состояние синапсов и свойства их деятельности в значительной мере определяются процессами, происходящими в соме клетки, в ее нейроплазме и ядре. Общеизвестна роль сомы в трофическом обеспечении передачи импульсов по путям их проведения и зависимости функционирования нейрона от его метаболизма. Это послужило основанием для поисков долгосрочного памятного следа в стойких изменениях химизма нейроплазмы.
Так как основным субстратом функциональных структур нейрона являются белковые соединения, а их различные модификации задаются матрицами РНК, на которых они синтезируются, то была выдвинута гипотеза о том, что информация, входящая в долгосрочную память, «записывается» структурой полинуклеотидной цепи молекулы (X. Хиден, 1959). По этой гипотезе разные условные потоки импульсов создают различное распределение ионных концентраций вдоль молекулы РНК и вызывают специфические для каждого сигнала перемещения нуклеотидов в их цепи. В результате каждый сигнал получает определенный отпечаток в структуре молекулы РНК. Поскольку структура молекулы РНК является матрицей для синтеза белковых соединений, специфические белки и медиаторы могут синтезироваться лишь по тому сигналу, который сформировал эту матрицу своим импульсным потоком. Все возможные перестановки и комбинации нуклеотидных элементов позволяют зафиксировать в молекулах РНК громадное количество информации. По приблизительному подсчету их информационная емкость достигает 1015–1020 бит, что перекрывает объем человеческой памяти.
В качестве экспериментального подтверждения гипотезы о переопределении нуклеотидов в молекуле РНК как механизме формирования долгосрочной памяти могут служить результаты опытов с крысами, которые обучались добираться к пище, балансируя на проволочном мостике (табл. 4).
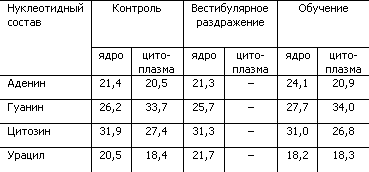
Таблица 4. Процентное содержание нуклеотидов в РНК ядра Дейтерса при выработке двигательного навыка у крыс (по X. Хидену, Е. Эльхази)
Как показывает табл. 4, у крыс, обучившихся сложным движениям, в нервных клетках ядра Дейтерса, ответственного за выполнение нового двигательного навыка, произошло изменение нуклеотидного состава РНК клеточного ядра. Увеличилась доля аденина, уменьшилась доля урацила. Простое вестибулярное раздражение не изменяло нуклеотидный состав. Исследование глиальных клеток ядра Дейтерса выявило сходные изменения РНК, что позволило сделать предположение об участии глии в кодировании поступающих сигналов путем изменения структуры нуклеотидных цепей РНК.
Однако гипотеза о нуклеотидном коде памятного следа встретилась со многими затруднениями, из которых главное — недолговечность молекул РНК. Выполняя информационные и транспортные функции, они могут сохранять свою структуру лишь в течение короткого времени, не соизмеримого с длительностью долгосрочной памяти. Эти и другие возражения ставят под сомнение РНК как носителя долгосрочного памятного следа, с чем, в сущности, согласился сам автор нуклеотидной гипотезы (X. Хиден, 1969).
В связи с этим внимание исследователей обратилось к более долговечным химическим компонентам структурно-функциональной организации нервной клетки. Памятный след стали искать в генетическом аппарате, который управляет деятельностью нервной клетки и определяет свойства на протяжении всей ее жизни. Наиболее стабильной структурой генетического аппарата являются молекулы ДНК, в которых кодируется наследственная информация. Логично было думать, что и информация, приобретаемая в течение жизни фиксируется этими структурами, специально приспособленными к хранению информационных программ. Поэтому были предприняты исследования ДНК как носителя долговременной памяти.
Анализ свойств ДНК и влияния ингибиторов ее синтеза на память привел к предположению, что формирование памятного следа связано с активацией определенных генов при деспирализации соответствующих участков цепи ДНК. Это происходит под влиянием ионных сдвигов, отражающих возбуждение нервной клетки при поступлении в нее сигналов.
На основании того, что снятие гистоновой блокады дерепрессирует гены, обусловливающие модификацию свойств клонов клеток была предложена нейроселективная теория долговременной памяти (В.В. Дергачев, 1967). Согласно этой теории при запоминании происходит селективная дерепрессия генов, реализующая определенную комбинацию из закодированных в геноме клонов нейроглиальных комплексов. В этих комплексах более лабильная функциональная ДНК программирует такое направление синтеза РНК и белков, которое определяет специфические свойства избирательной возбудимости нервных клеток. О значении ДНК в формировании долгосрочной памяти свидетельствуют опыты, показывающие нарушения памяти при действии ингибиторов синтеза рибонуклеиновых кислот даже в то время, когда синтез белков продолжался.
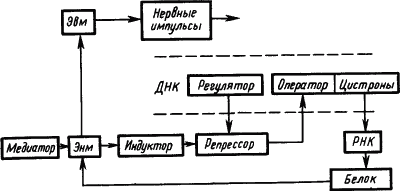
Электрофизиологические исследования механизмов привыкания нервных клеток к действию раздражителей привели к представлению, что памятный след отражает не столько временные, сколько пространственные характеристики сигналов. Он представляет собой не «паттерн импульсов», а «паттерн синапсов», кодирующих своим расположением и состоянием значение сигнала (Е.Н. Соколов, 1969). При этом медиаторы через рецепторы синаптических мембран вызывают активацию одних и инактивацию других оперонов молекулы ДНК. В результате формируется система обеспечения синтеза специфических белков, реализующих функцию «обученной нервной клетки (рис. 25).
Рис. 25. Участие функциональных структур генетического аппарата молекулы ДНК в долговременной памяти (по Е.Н. Соколову):
пунктиром обозначен участок молекулы ДНК; Эвм — электровозбудимая мембрана, Энм — электроневозбудимая мембрана
За последнее время получены новые доказательства участия ДНК в механизмах долговременной памяти. Показано, что при выработке условных рефлексов происходят своеобразные изменения структуры молекулы ДНК в нервных клетках мозга крыс. Эти изменения состояли в увеличении степени метилирования ДНК, повышающего активность определенных генов. Они были четко выражены в коре больших полушарий и гиппокампе и оказались незначительными в мозжечке. При такой «мягкой» обратимой модификации ДНК ее нуклеотидный состав оставался неизменным (табл. 5).