Вопросы
1. Уровни построения движений в нервной системе человека.
2. Спинной мозг – рефлекторный уровень построения движений.
3. Строение серого вещества спинного мозга.
4. Рефлексы мозгового ствола.
5. Нейрофизиология глазодвигательных реакций.
Литература
Бернштейн Н. А. О построении движений. М.: Изд. АН СССР, 1970.
Магнус Р. Установка тела. М.; Изд. АН СССР, 1962.
Шульговский В. В. Физиология целенаправленного поведения млекопитающих. М.: Изд-во Моск. ун-та, 1993.
ФИЗИОЛОГИЯ МОЗЖЕЧКА
Мозжечок, или малый мозг, представляет собой надсегментарную структуру, расположенную над продолговатым мозгом и мостом, позади больших полушарий мозга. Мозжечок состоит из нескольких частей, различных по происхождению в эволюции позвоночных животных.
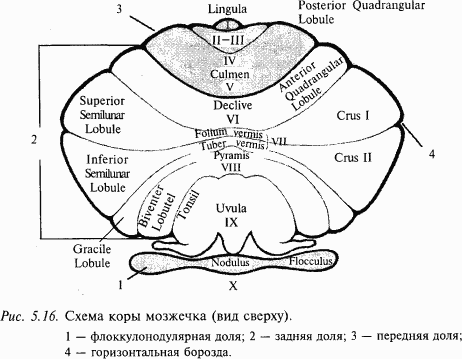
У человека мозжечок состоит из двух полушарий, находящихся по бокам от червя. К филогенетически более древней части мозжечка млекопитающих относят переднюю долю и флоккулонодулярную часть задней доли. Эти структуры мозжечка преимущественно связаны со спинным мозгом и вестибулярным аппаратом, тогда как полушария в основном получают информацию от мышечных и суставных рецепторов, а также от зрительного и слухового анализаторов. На рис. 5.16 представлена схема мозжечка млекопитающего (см. приложение 6), отражающая плотность вестибулярных, проприоцептивных (от мышц, сухожилий и суставов) и корковых афферентных проекций в различные зоны мозжечка. Согласно этой классификации кора мозжечка делится на три области:
1) архицеребеллум (старый мозжечок) – флоккулонодулярная доля (долька X); в ней оканчиваются преимущественно вестибулярные афференты и волокна от вестибулярных ядер; вестибулярные волокна проецируются также частично в язычок (lingula – долька I) и каудальную часть втулочки (uvula – долька IX), которые обычно относят также к архицеребеллуму;
2) палеоцеребеллум (древний мозжечок) включает переднюю долю (дольки II–V), простую дольку (долька VI) и заднюю часть корпуса мозжечка (дольки VIII–IX); палеоцеребеллум тесно связан со спинным мозгом, а также имеет двусторонние связи с сенсомоторной областью коры больших полушарий;
3) неоцеребеллум (новый мозжечок) включает среднюю часть корпуса мозжечка (долька VII и частично дольки VI и VIII), которая получает информацию от коры больших полушарий, а также от слуховых и зрительных рецепторов. Обратите внимание, что основная часть полушарий мозжечка принадлежит новому мозжечку, который лучше всего развит у человека.
В толще мозжечка находятся три пары ядер: зубчатое, расположенное латерально; ядро шатра – медиально; пробковидное и округлое ядра – между ними.
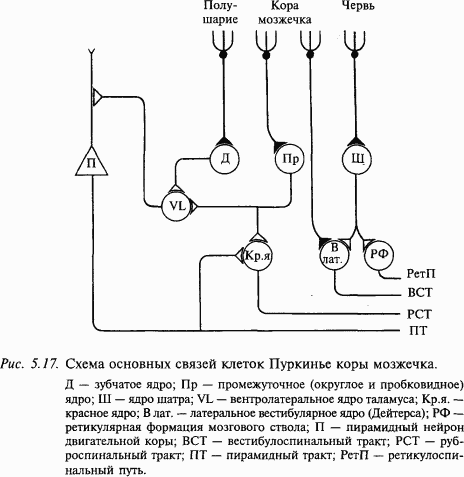
Единственным эфферентным выходом из коры мозжечка являются аксоны клеток Пуркинье, образующие синапсы с нейронами внутримозжечковых ядер и нейронами латерального вестибулярного ядра (рис. 5.17). Тесная связь ядра Дейтерса с корой мозжечка дает основание рассматривать его функционально как внутримозжечковое ядро. Все остальные образования головного и спинного мозга не получают прямых эфферентов из коры мозжечка. Ядра шатра посылают волокна к ядрам Дейтерса и к ретикулярной формации продолговатого мозга. Из области ретикулярной формации, где оканчиваются пути от мозжечка, берет начало ретикулоспинальный путь. Промежуточные ядра посылают аксоны в средний мозг, в том числе к красному ядру. Мощные пучки волокон, образованные преимущественно аксонами нейронов зубчатого ядра, направляются к вентролатеральному ядру таламуса, где происходит синаптическое переключение, и аксоны постсинаптических нейронов идут в моторную область коры больших полушарий; часть аксонов направляется к базальным ядрам. Таким образом, мозжечок не имеет самостоятельных двигательных систем, но образует обширные связи со всеми моторными системами: кортикоспинальной (пирамидной), руброспинальной, ретикулоспинальной, вестибулоспинальной, а также с полосатым телом.
Афферентные и эфферентные волокна мозжечка собраны в три пары массивных волокнистых пучков, известных как мозжечковые ножки. Афферентные волокна входят в мозжечок в основном через нижние и средние ножки мозжечка. Эфферентные волокна проходят преимущественно через верхние ножки. Однако имеются исключения: некоторая часть спинно-мозжечковых путей входит через верхние ножки, а некоторые эфферентные волокна от флоккулонодулярной доли и ядра шатра проходят через нижние ножки.
Ядра шатра направляют волокна через нижние ножки к вестибулярным ядрам и ретикулярной формации продолговатого мозга и моста. Промежуточные и зубчатые ядра посылают волокна через верхние ножки преимущественно к среднему мозгу и таламусу, особенно к красному ядру. Основная часть церебелло-таламических волокон отходит от зубчатых ядер. Эти волокна проецируются также к красному ядру, полосатому телу. Таким образом, влияния мозжечка на спинальные мотонейроны осуществляются через вестибулоспинальные и ретикулоспинальные пути, а на прецентральную область коры – через вентролатеральное ядро таламуса.
Строение коры мозжечка. Кора мозжечка различных представителей позвоночных, включая человека, построена по единому плану и состоит из трех слоев (см. приложение 6). Поверхностный, или молекулярный, слой содержит разветвления дендритов клеток Пуркинье и параллельные волокна. Клетки Пуркинье имеют уплощенный дендрит, ориентированный параллельно сагиттальным зонам долек (folia) мозжечка. Дендриты и аксоны звездчатых клеток в молекулярном слое расположены таким же образом, тогда как параллельные волокна ориентированы строго трансверзально (перпендикулярно) по отношению к фоллиуму и сагиттальному направлению мшистых волокон. У кошки, например, параллельные волокна имеют среднюю длину 2 мм (диапазон до 5–7 мм). Одно параллельное волокно пересекает около 700 дендритов клеток Пуркинье.
Проекции мшистых и лазающих волокон в коре мозжечка организованы в виде сагиттальных полосок. В нижней части молекулярного слоя находятся также тела корзинчатых клеток, аксоны которых идут перпендикулярно направлению листка коры мозжечка и оплетают тела и начальные сегменты аксонов клеток Пуркинье. В молекулярном слое имеется также небольшое число звездчатых клеток.
Самый нижний, гранулярный слой содержит клетки-зерна, или гранулярные клетки. От тела клетки-зерна отходит 4–7 коротких дендритных отростков, с которыми мшистые волокна образуют синапсы. Аксон клетки-зерна поднимается вертикально вверх и в молекулярном слое Т-образно ветвится, образуя параллельные волокна. Плоскость его ветвления перпендикулярна плоскости ветвления дендритов клеток Пуркинье. В гранулярном слое находятся клетки Гольджи, аксоны которых восходят в молекулярный слой. Ганглиозный слой находится между описанными выше молекулярным и гранулярным слоями и содержит тела клеток Пуркинье.
Афферентный вход в кору мозжечка осуществляется в основном через две системы волокон: лазающие и моховидные, или мшистые. Каждая клетка Пуркинье получает вход только от одного лазающего волокна (медиатор – аспартат), которое заканчивается на начальной (проксимальной) части дендрита. Лазающие волокна представляют собой аксоны нейронов, тела которых находятся в нижних оливах. По этому входу оказывается мощное возбуждающее действие: клетка Пуркинье деполяризуется на 10–15 мс, и в ней развиваются кальцийзависимые потенциалы действия. За потенциалом действия наступает следовая деполяризация, которая возникает вследствие активации кальцийзависимой калиевой проводимости соматической мембраны. После следовой деполяризации наступает следовая гиперполяризация. По этой причине клетка Пуркинье может возбуждаться по этому входу не более 1–2 раз в 1 с.