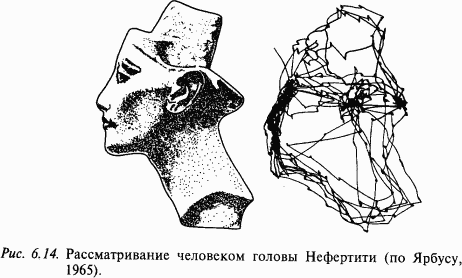
Движения глаз и зрительное восприятие. При рассматривании различных объектов внешнего мира глаза совершают быстрые (саккады) и медленные следящие движения. Благодаря медленным следящим движениям изображение движущихся объектов удерживается на fovea. При рассматривании хорошо структурированного изображения глаза совершают саккады, перемежающиеся с фиксацией взора. Если человек рассматривает изображение в течение некоторого времени, то запись перемещений глаза воспроизводит достаточно грубо контур и наиболее информативные детали рассматриваемого объекта. Например, при рассматривании лица особенно часто фиксируются рот и глаза (рис. 6.14). Специальные эксперименты показали, что во время саккады зрительное восприятие блокируется. Можно предложить несколько механизмов этого феномена. Предполагают, что во время саккады по сильно структурированному фону флюктуации интенсивности в каждой точке превышают частоту слияния мельканий. Другой механизм, блокирующий зрительное восприятие во время саккады, –центральное торможение. Когда движущийся объект появляется на периферии зрительного поля, он вызывает рефлекторную саккаду, которая может сопровождаться движением головы. Основой нейрофизиологического механизма этого рефлекса являются детекторы движения в зрительной системе. Биологически рефлекс оправдан тем, что благодаря ему внимание переключается на новый объект, появившийся в поле зрения.
Вопросы
1. Строение глаза.
2. Схема зрительной системы человека.
3. Строение рецептивных полей нейронов сетчатки, латерального коленчатого тела и коры.
4. Обработка зрительных сигналов в ассоциативных полях коры.
5. Роль движения глаз в зрительном восприятии.
Литература
Подвигин Н. Ф. Динамические свойства нейронных структур зрительной системы. Л.: Наука, 1979.
Супин А. Я. Нейрофизиология зрения млекопитающих. М.: Наука, 1981.
Хьюбель Д. Глаз, мозг, зрение. М.: Мир, 1990.
Шевелев И. А. Нейроны зрительной коры. Адаптивность и динамика рецептивных полей. М.: Наука, 1981.
НЕЙРОФИЗИОЛОГИЯ СЛУХОВОЙ СИСТЕМЫ
Мир, окружающий человека, полон звуков. Правда, человек слышит не все звуки этого мира, а только те, которые находятся в диапазоне от 20 до 20 000 Гц. Известно, что многие животные обладают значительно более широким диапазоном слышимых звуков. Например, дельфины «слышат» звуки частотой до 170 000 Гц, собаки и кошки также имеют более широкий диапазон слышимых звуков. Но слуховая система человека предназначена преимущественно для того, чтобы слышать речь другого человека, и в этом отношении ее совершенство нельзя даже близко сравнивать со слуховыми системами других млекопитающих.
Наружное ухо включает ушную раковину и наружный слуховой проход. Рельеф ушной раковины играет значительную роль в восприятии звуков. Если, например, этот рельеф уничтожить, залив воском, человек заметно хуже определяет направление источника звука. Наружный слуховой проход человека в среднем имеет длину около 9 см. Есть данные, что трубка такой длины и схожего диаметра имеет резонанс на частоте около 1 кГц, другими словами, звуки этой частоты немного усиливаются. Среднее ухо отделено от наружного барабанной перепонкой, которая имеет вид конуса с вершиной, обращенной в барабанную полость.
Среднее ухо человека состоит из барабанной полости, которая соединена евстахиевой трубой с полостью глотки, и барабанной перепонки с тремя слуховыми косточками (молоточком, наковальней и стремечком), сочлененных между собой (рис. 6.15). Слуховые косточки действуют как рычаги, улучшая передачу звуковых колебаний из воздушной среды наружного слухового прохода в более плотную водную среду внутреннего уха. Здесь имеются также две маленькие мышцы – одна прикреплена к ручке молоточка, а другая – к стремечку (на рис. 6.15 не показаны). Сокращение этих мышц предотвращает слишком большие колебания косточек, вызванных громкими звуками. Это так называемый акустический рефлекс. Таким образом, основной функцией акустического рефлекса является защита улитки от повреждающей стимуляции.
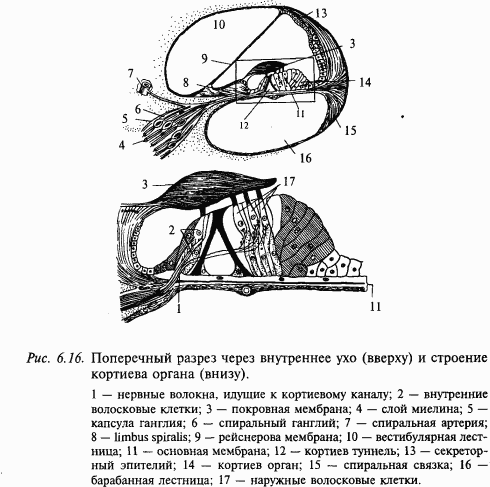
Внутреннее ухо включает два рецепторных аппарата: вестибулярный и слуховой. Вестибулярный аппарат был уже рассмотрен выше. Опишем слуховой орган. Костный канал улитки разделен двумя мембранами – основной, или базилярной, и рейснеровой – на три отдельных канала, или лестницы: барабанную, вестибулярную и среднюю. Каналы внутреннего уха заполнены жидкостями, ионный состав которых в каждом канале специфичен. Средняя лестница заполнена эндолимфой с высоким содержанием ионов калия; она сообщается с эндолимфой вестибулярного органа. Две другие лестницы заполнены перилимфой, состав которой не отличается от тканевой жидкости. Внутри лестницы имеется еще один канал – кортиев туннель. По обе стороны от него находятся опорные клетки и ряды волосковых клеток (слуховые рецепторы), называемые соответственно внутренними и наружными клетками. Волосковые и опорные клетки, а также кортиев туннель покрыты соединительнотканной покровной (текториальной) мембраной, играющей важную роль в возбуждении слуховых рецепторов.
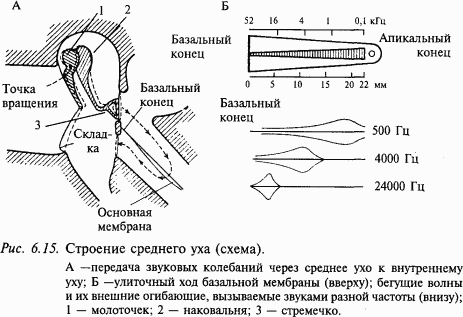
Эта структура (нейроэпителий и покровная мембрана), протянувшаяся по всей длине улитки, называется кортиевым органом (рис. 6.16). Вестибулярная и барабанная лестницы на вершине улитки соединяются через небольшое отверстие – геликотрему, средняя лестница заканчивается слепо (см. рис. 6.15).
Средняя лестница, заполненная эндолимфой, заряжена положительно (до +80 мВ) относительно двух других лестниц. Если учесть, что потенциал покоя отдельных волосковых клеток около – 80 мВ, то в целом разность потенциала (эндокохлеарный потенциал) на участке средняя лестница – кортиев орган может составить около 160 мВ. Эндокохлеарный потенциал улитки чрезвычайно чувствителен к недостатку кислорода. Источником эндокохлеарного потенциала является, по-видимому, сосудистая полоска. Эндокохлеарный потенциал играет важную роль в возбуждении волосковых клеток. Предполагают, что волосковые клетки поляризованы этим потенциалом до критического уровня. В этих условиях минимальные механические воздействия могут вызвать возбуждение рецептора.
Звуковая волна действует на барабанную перепонку, и далее через систему косточек звуковое давление передается на овальное окно и воздействует на перилимфу вестибулярной лестницы (см. рис. 6.15). Поскольку жидкость несжимаема, перемещение перилимфы может передаваться через геликотрему в барабанную лестницу, а оттуда через круглое окно – обратно в полость среднего уха. Перилимфа может перемещаться и более коротким путем: рейснерова мембрана изгибается, и через среднюю лестницу давление передается на основную мембрану, затем в барабанную лестницу и через круглое окно в полость среднего уха. Именно в последнем случае раздражаются слуховые рецепторы. Колебания основной мембраны приводят к смещению волосковых клеток относительно покровной мембраны и, следовательно, к их деполяризации и далее к возбуждению волокон слухового нерва.
Венгерский ученый Г. Бекеши (1951) предложил «теорию бегущей волны», позволяющую понять, как звуковая волна определенной частоты возбуждает волосковые клетки, находящиеся в определенном месте основной мембраны. Эта теория получила всеобщее признание. Основная мембрана расширяется от основания улитки к ее вершине примерно в 10 раз (у человека от 0,04 до 0,5 мм). Предполагается, что основная мембрана закреплена только по одному краю, остальная ее часть свободно скользит, что соответствует морфологическим данным. Теория Бекеши объясняет механизм анализа звуковой волны следующим образом: высокочастотные колебания проходят по мембране лишь короткое расстояние, а длинные волны распространяются далеко. Тогда начальная часть основной мембраны служит высокочастотным фильтром, а длинные волны проходят весь путь до геликотремы. Соединив все точки максимального перемещения колеблющейся мембраны, можно получить воображаемую поверхность, которую называют огибающей колебания. На рис. 6.15 показаны в поперечном разрезе огибающие колебаний основной мембраны для разных частот. Чистые тоны имеют огибающие с одним максимумом, от которого пологий спуск ведет к основанию, а более крутой – к верхушке улитки. Максимальные перемещения для разных частот происходят в разных точках основной мембраны: чем ниже тон, тем ближе его максимум к верхушке улитки. Таким образом, высота звука кодируется местом на основной мембране.