8. Оптический установочный рефлекс играет особенно важную роль в правильной установке головы у высших млекопитающих. Например, обезьяна с разрушенным лабиринтом может правильно устанавливать голову, но если надеть ей колпак, закрывающий глаза, т.е. лишить зрения, она уже не сможет правильно устанавливать голову в пространстве.
Таким образом, в организме животных взаимодействует ряд установочных рефлексов, благодаря которым обеспечивается нормальное положение головы и тела. Описанные установочные рефлексы осуществляются нервными центрами на уровне не выше среднего мозга.
9. Лифтная реакция представляет собой рефлекс на прямолинейное ускорение. Рефлекс состоит в том, что при поднятии площадки, на которой находится животное, тонус экстензоров снижается, и конечности подгибаются; при опускании площадки развивается противоположная реакция: экстензоры активируются и конечности разгибаются. Эта реакция легко вызывается и у человека при подъеме или опускании в скоростном лифте.

Описанные выше установочные рефлексы легко наблюдать на интактной кошке, которая свободно падает из положения на спине (рис. 5.14). Прежде всего, активируются лабиринтные рефлексы на голову, благодаря которым она поворачивается по направлению к нормальному положению. К этой реакции присоединяется шейный установочный рефлекс, вследствие чего за головой следует тело, сначала грудной отдел и только затем таз. Происходит своего рода штопорообразное движение животного в пространстве, начинающееся с головы. При этом передние конечности разгибаются. Дальнейшее прямолинейное смещение головы в пространстве при свободном падении вызывает разгибание задних конечностей. Последнее можно объяснить лабиринтным рефлексом на падение (линейное ускорение). Благодаря совокупному действию этих рефлексов голова и тело кошки принимают нормальное положение, конечности тонически разгибаются и готовы принять вес тела, когда животное достигнет земли.
Таким образом, положение тела и головы в пространстве и по отношению друг к другу принадлежит к наиболее застрахованным и автоматически выполняемым реакциям ЦНС. Благодаря взаимной деятельности большого количества разнородных рефлексов осуществляется единая конечная цель – правильная установка головы и тела в пространстве. Статокинетические и статические реакции взаимно дополняют друг друга: благодаря кинетическим реакциям выполняется движение, приводящее отдельные части тела в такое положение, в котором они потом удерживаются статическими рефлексами.
Глазодвигательная система млекопитающих. В эволюции наземных позвоночных животных возникла необходимость сохранять неподвижность изображения на сетчатке не только в состоянии покоя или медленных движений, но также во время локомоции. Эволюция глазодвигательной системы у этих животных происходила в тесном взаимодействии с эволюцией зрительной системы. С точки зрения эволюции глазодвигательной системы всех позвоночных животных можно условно разделить на две группы: «фовеальные» животные, сетчатка которых имеет fovea – и они обладают достаточно острым зрением, и «афовеальные» животные – с недостаточно острым зрением. У «фовеальных» животных (к ним относится и человек) глаза расположены во фронтальной плоскости, у них есть бинокулярное зрение. Зрительная ямка, или fovea, – это небольшая область в сетчатке с повышенной плотностью фоторецепторов, причем только в этой области находятся колбочки – фоторецепторы дневного (и цветового) зрения. Глазодвигательная система млекопитающих, которая сформировалась как необходимое дополнение зрительной системы, позволяет рассматривать зрительный мир. С точки зрения физиологии непрерывно решается задача – движением глаз поместить на зрительную ямку наиболее значимые части изображения. Что будет, если отключить fovea? Это происходит с каждым из нас при снижении уровня освещенности, например, вечером. Известно, что мы в этих условиях не видим мелких предметов, не можем читать. Для такого зрения движения глаз не обязательны. Действительно, у животных с невыраженной fovea, например собаки, коровы, быстрые движения глаз отсутствуют.
Глазодвигательная система млекопитающих выполняет следующие задачи: 1) сохраняет неподвижным изображение внешнего мира на сетчатке во время движения животного относительно этого мира; 2) выделяет во внешнем мире некоторые объекты, помещает их в зоне сетчатки с высоким разрешением (например, fovea у человека) и прослеживает их движениями глаз и головы; 3) осуществляет скачкообразные (саккадические) перемещения взора для сканирования (рассматривания) внешнего мира. «Афовеальные» животные, например кошка, собака, кролик и др., не заботятся о положении изображения на сетчатке, и вестибулярная и оптокинетическая системы только предотвращают скольжение изображения по сетчатке. У человека в связи с изложенными выше требованиями, кроме вестибулоокулярной и оптокинетической систем, имеется также система прослеживания, вергенции и саккад. У него развились также рефлекторные механизмы содружественного движения глаз и головы.
Саккадная система в полной мере развита только у обезьян и человека. Именно у них есть бинокулярное зрение, и есть «фовеальное» зрение.
Саккады – это быстрые содружественные отклонения глаз в быструю фазу вестибулярного и оптомоторного нистагмов, начальная фаза реакции прослеживания, когда скачком глаза «захватывается» движущаяся зрительная цель при зрительном обследовании внешнего мира. Структуры мозга, обеспечивающие быстрые, скачкообразные движения глаз – саккады, находятся в ретикулярной формации мозгового ствола. Однако высший центр произвольного управления движениями глаз локализован в коре лобной доли мозга и называется фронтальным глазодвигательным полем (8-е и 10-е поля).
Активность нейронов мозгового ствола. Схема, представленная на рис. 5.15, показывает связи и вероятные события, приводящие к саккаде глаза. Важными элементами этой схемы являются пачечные нейроны и нейроны-паузы. Во время фиксации взора клетки-паузы разряжаются тонически с высокой частотой; так как тормозные связи от них идут к тормозным и возбудительным пачечным нейронам, это должно предотвращать формирование пачки импульсов. Предполагают, что возбудительные пачечные нейроны проецируются к тоническим нейронам и происходит интегрирование их активности, в результате чего образуется сигнал, необходимый для поддержания глаза в эксцентричном положении (так называемый нейронный интегратор). В действительности нейронный интегратор, по-видимому, имеет большую протяженность. Установлено, что разрушение в области мозгового ствола и мозжечка приводит к параличу взора, т.е. неспособности поддерживать эксцентричное положение взора.
По логике схемы стволового генератора запускающий сигнал формируется клетками-паузами. Установлено, что они получают афферентацию из средних и глубоких слоев верхнего двухолмия. Электрофизиологическими методами также было показано, что клетки-паузы могут возбуждаться моносинаптически на электрическую стимуляцию нижних слоев верхнего двухолмия. Таким образом, нейронная схема генератора саккады «работает» по принципу «спускового крючка» – клетка-пауза постоянно тормозит пачечные нейроны. Как только из верхнего двухолмия затормаживается разряд клетки-паузы, в этот интервал растормаживается пачечный нейрон и этот сигнал подается на соответствующие мотонейроны, что и приводит к саккаде.
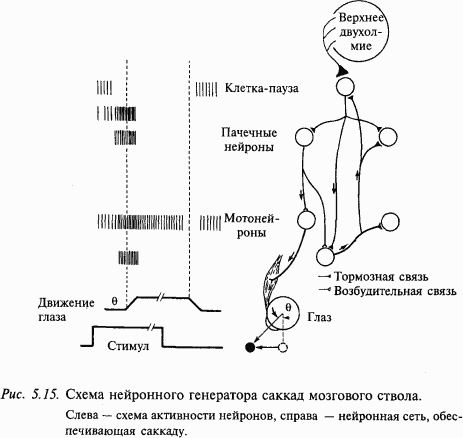
На основании электрофизиологических данных было высказано предположение, что пачечные нейроны со спонтанным разрядом являются промежуточным звеном между нижними слоями переднего двухолмия и парамедианной ретикулярной формацией моста. По мнению исследователей, эти нейроны могут получать конвергентные входы из ряда источников, в том числе из фронтального глазодвигательного поля. Известно, что дефицит саккад возникает только при одновременном разрушении переднего двухолмия и фронтального глазодвигательного поля.