Х. п. могут возникать спонтанно, но частота их резко возрастает под влиянием физических и химических факторов (см. Мутагены , Мутагенез ). Небольшие делеции и дупликации могут быть следствием неравного кроссинговера. Х. п. играют большую роль в эволюции организмов: дупликации представляют главный источник увеличения числа генов; инверсии и транслокации могут вести к генетической изоляции гомозиготных по ним особей, более плодовитых, чем гетерозиготы. При всех Х. п. иногда наблюдается т. н. эффект положения гена , заключающийся в том, что ген, перенесённый в новое место хромосомы, изменяет своё действие на фенотип организма. Х. п. могут быть использованы в практических целях для изменения групп сцепления генов , определяющих хозяйственно ценные признаки организмов.
С. М. Гершензон.
Хромосомный набор
Хромосо'мный набо'р, совокупность хромосом , заключённая в ядре любой клетки тела растительного или животного организма; характеризуется постоянным для каждого биологического вида числом хромосом, определённой их величиной и морфологическими особенностями. См. Кариотип .
Хромосомы
Хромосо'мы (от хромо... и сома ), органоиды клеточного ядра, совокупность которых определяет основные наследственные свойства клеток и организмов. Полный набор Х. в клетке, характерный для данного организма, называется кариотипом . В любой клетке тела большинства животных и растений каждая Х. представлена дважды: одна из них получена от отца, другая — от матери при слиянии ядер половых клеток в процессе оплодотворения. Такие Х. называются гомологичными, набор гомологичных Х. — диплоидным. В хромосомном наборе клеток раздельнополых организмов присутствует пара (или несколько пар) половых хромосом, как правило, различающихся у разных полов по морфологическим признакам; остальные Х. называются аутосомами. У млекопитающих в половых Х. локализованы гены , определяющие пол организма; у плодовой мушки дрозофилы пол определяется соотношением половых хромосом и аутосом (балансовая теория определения пола).
Первоначально Х. были описаны как интенсивно окрашивающиеся основными красителями плотные тельца (немецкий учёный В. Вальдейер, 1888). Однако оказалось, что внешний вид Х. существенно меняется на разных стадиях клеточного цикла, и как компактные образования с характерной морфологией Х. четко различимы в световом микроскопе лишь в период клеточного деления — в метафазе митоза и мейоза (рис. 1 , 2 ). Основу Х. на всех стадиях клеточного цикла составляют хромонемы — нитевидные структуры, которые во время деления клетки плотно закручены, обусловливая спирализацию хромосом , а в неделящейся клетке раскручены (деспирализованы). При завершении деления клетки разошедшиеся к её полюсам Х. разрыхляются и окружаются ядерной мембраной. В период между двумя делениями клетки (эта стадия клеточного цикла называется интерфазой ) деспирализация Х. продолжается и они становятся малодоступными для наблюдения в световой микроскоп. Морфология Х. эукариот существенно отличается от таковой у прокариот и вирусов. Прокариоты (доядерные) и вирусы содержат обычно одну линейную или кольцевую Х., которая не имеет надмолекулярной укладки и не отделена от цитоплазмы ядерной оболочкой. Понятие Х. к генетическому аппарату прокариот применимо лишь условно, т.к. оно сформировалось при изучении Х. эукариот и подразумевает наличие в Х. не только сложного комплекса биополимеров (нуклеиновых кислот и белков), но и специфической надмолекулярной структуры. Поэтому ниже даётся описание только Х. эукариот. Изменения внешнего вида Х. в клеточном и жизненном циклах обусловлены особенностями функционирования Х. Общий же принцип их организации, индивидуальность и непрерывность Х. в ряду клеточных поколений и организмов сохраняются неизменными. Доказательства тому получены при биохимическом, цитологическом и генетическом исследованиях Х. разных организмов. Они легли в основу хромосомной теории наследственности .
Молекулярные основы строения Х. Значение Х. как клеточных органоидов, ответственных за хранение, воспроизведение и реализацию наследственной информации, определяется свойствами биополимеров, входящих в их состав. Первая молекулярная модель Х. была предложена в 1928 Н. К. Кольцовым , предугадавшим принципы их организации. Запись наследственной информации в Х. обеспечивается строением молекулы дезоксирибонуклеиновой кислоты (ДНК), её генетическим кодом . В Х. сосредоточено около 99% всей ДНК клетки, остальная часть ДНК находится в других клеточных органоидах, определяя цитоплазматическую наследственность. ДНК в Х. эукариот находится в комплексе с основными белками — гистонами и с негистоновыми белками, которые обеспечивают сложную упаковку ДНК в Х. и регуляцию её способности к синтезу рибонуклеиновых кислот (РНК) — транскрипции .
Х. в интерфазе. Х. выполняет свои основные функции — репродукцию и транскрипцию — в интерфазе, поэтому строение Х. на этой стадии клеточного цикла представляет особый интерес. В интерфазе Х. плохо различимы потому, что в связи с активным синтезом РНК многие участки Х. (т. н. эухроматин ) сильно раскручены; другие же (гетерохроматин ) не участвуют в синтезе РНК и продолжают сохранять плотную упаковку (см. также Хромоцентр ). В эухроматиновых участках, помимо элементарных дезоксирибонуклеопротеидных нитей (ДНП), имеются рибонуклеопротеидные частицы диаметром 200—500
, называемые РНП-гранулами, интергранулами и перихроматиновыми гранулами. Эти частицы представляют собой форму упаковки РНК, синтезированной на Х. и соединённой с белком, и служат для завершения образования информационной РНК и переноса её в цитоплазму.
Для изучения интерфазных Х. используют либо биохимические методы выделения вещества Х. — хроматина и разделения его на эухроматин и гетерохроматин, либо электронно-микроскопическое исследование интактных ядер и изолированного хроматина; как модели интерфазных Х. используют гигантские Х. типа ламповых щёток из ооцитов животных и многонитчатые (политенные) Х. двукрылых. В Х. типа ламповых щёток неактивные участки имеют вид плотно упакованных структур — хромомер (рис. 2 , 3 ), которые обнаруживаются и в Х. соматических клеток, особенно в профазе митоза, и рассматриваются как характерные морфологические, а возможно и функциональные, единицы Х. В участках Х., активно синтезирующих РНК, хромомеры раскручиваются и образуют боковые петли, в которых молекулы РНК, соединяясь с белком, образуют рибонуклеопротеиды (РНП) — частицы, представляющие собой форму упаковки генных продуктов и различающиеся в отдельных боковых петлях по размерам и морфологическим признакам. В политенных Х., возникающих в тканях двукрылых и некоторых растений за счёт многократной репликации (удвоения) исходной Х. без последующего расхождения дочерних Х., неактивные участки имеют форму дисков, а активные образуют вздутия — пуфы . В пуфах, так же как и в Х. типа ламповых щёток, содержатся частицы РНП диаметром 200—500
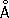
. Электронно-микроскопические и биохимические исследования показали, что и в хроматине, выделенном из клеток, и в интактных ядрах, и в гигантских Х. основной структурной единицей является дезоксирибонуклеопротеидная нить (ДНП) диаметром,100—200
.
Изучение политенных Х. в разных тканях и на разных стадиях развития двукрылых показало, что число и набор активных пуфов имеют тканевую и видовую специфичность. Это значит, что хотя все клетки многоклеточного организма имеют одинаковый набор генов, линейно расположенных в каждой Х., набор активных и неактивных в синтезе РНК участков Х. различается в каждом типе клеток и на разных стадиях развития, т. е. один и тот же участок находится в одних тканях в эухроматическом, в других — в гетерохроматическом состоянии. Отдельные участки Х. находятся в гетерохроматическом состоянии в интерфазе разных типов клеток; как правило, они отличаются присутствием высокоповторяющихся последовательностей ДНК. Постоянно функционирующим в интерфазе всех типов клеток является ядрышковый организатор — участок Х., где сосредоточены гены рибосомной РНК. В этой области формируется ядрышко , которое долго считали самостоятельным органоидом клетки. Оно является местом формирования предшественников рибосом .