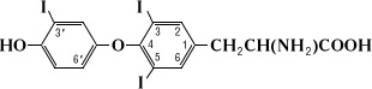
Секретируется в кровь в значительно меньших количествах, чем тироксин, но оказывает более быстрое действие на обмен веществ, так как менее прочно связан со специфическими белками плазмы и потому легче проникает в тканевую жидкость и клетки.
Трикала
Три'кала (Trikkala), город в Центральной Греции, в Фессалии. Административный центр нома Трикала. 34,8 тыс. жителей (1971). Текстильная, пищевая, табачная, кожевенно-обувная, дерево- и металлообрабатывающая промышленность.
Трикальцийфосфат
Трикальцийфосфа'т , Ca3 (PO4 )2 , соль ортофосфорной кислоты; см. Фосфаты кальция .
Трикар Жан
Трика'р (Tricart) Жан (р. 16.9.1920, Монморанси), французский геолог и геоморфолог. После окончания историко-литературного факультета в Парижском университете работал ассистентом там же (1945—48). С 1947 сотрудник службы геологических карт Франции. Профессор (с 1955) и директор Центра прикладной географии Страсбурского университета (с 1957). Президент комиссии прикладной геоморфологии Международного географического союза (с 1955). Основные работы по структурной и климатической геоморфологии, по прикладным и методологическим вопросам геоморфологии. Изучал геологическое строение дельты Сенегала, Среднего Нигера, бассейна Бразилии, Венесуэлы; водные ресурсы Перу, Чили, различных бассейнов Франции. Т. — основатель и директор «Revue de géomorphologique dynamique».
Трикарбоновых кислот цикл
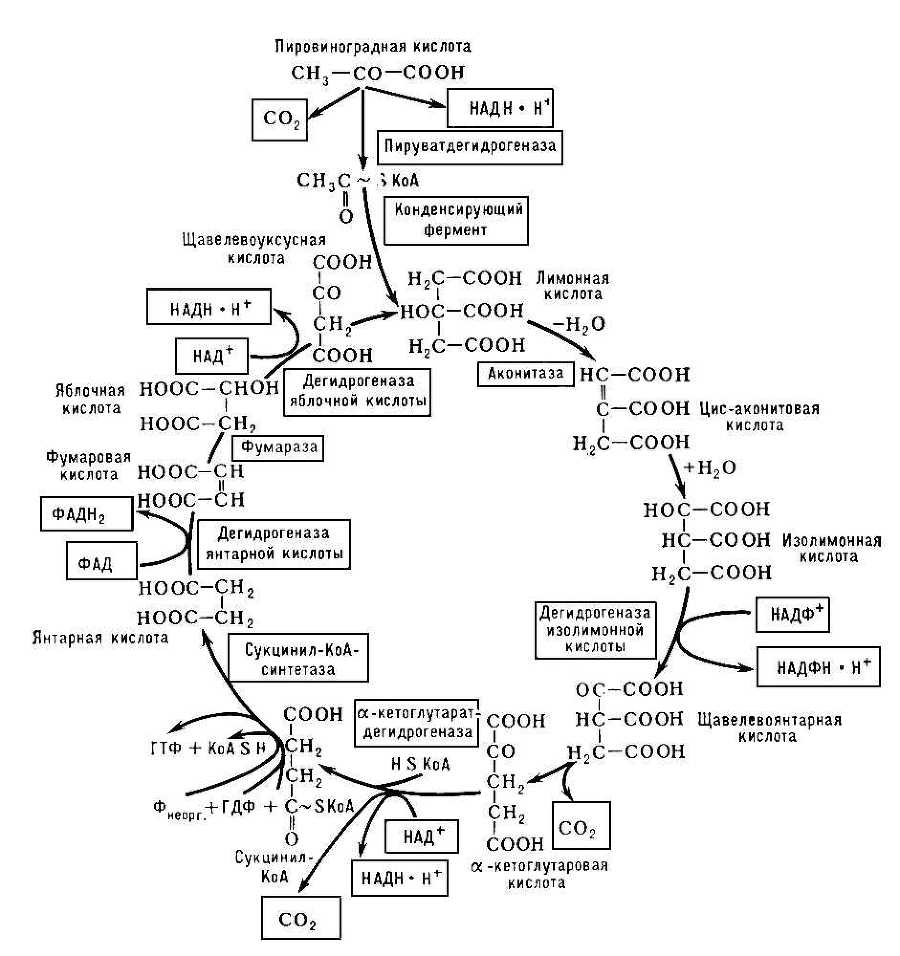
Трикарбо'новых кисло'т цикл , цикл лимонной кислоты, цикл Кребса, широко представленный в организмах животных, растений и микробов путь окислительных превращений ди- и трикарбоновых кислот, образующихся в качестве промежуточных продуктов при распаде белков, жиров и углеводов. Открыт Х. Кребсом и У. Джонсоном (1937). Т. к. ц., локализованный в митохондриях , начинается с лимонной кислоты и заканчивается образованием щавелевоуксусной кислоты, CO2 и восстановлением коферментов дегидрогеназ: никотинамидадениндинуклеотида (НАД) и флавинадениндинуклеотида (ФАД). К субстратам Т. к. ц. относятся трикарбоновые кислоты — лимонная, цис-аконитовая, изолимонная, щавелевоянтарная и дикарбоновые кислоты — кетоглутаровая, янтарная, фумаровая, яблочная и щавелевоуксусная. К субстратам Т. к. ц. следует отнести и уксусную кислоту, которая в активной форме, то есть в виде ацетилкофермента А (ацетил-КоА), участвует в конденсации с щавелевоуксусной кислотой, приводящей к образованию лимонной кислоты. Именно ацетильный остаток, вошедший в структуру лимонной кислоты, подвергается окислению; атомы углерода окисляются до CO2 , атомы водорода частично акцептируются коферментами дегидрогеназ, частично в протонированной форме переходят в раствор, то есть в окружающую среду.
Обычно указывают на пировиноградную кислоту (пируват), образующуюся при гликолизе в реакциях переаминирования и занимающую одно из центральных мест в перекрещивающихся путях обмена веществ, как на исходное соединение для образования ацетил-КоА. Действительно, под влиянием фермента сложной структуры — пируватдегидрогеназы — осуществляется окисление пирувата с образованием CO2 (первое декарбоксилирование), ацетил-КоА и происходит восстановление НАД (см. схему ). Однако окисление пирувата далеко не единственный путь образования ацетил-КоА, который является характерным продуктом митохондриального окисления жирных кислот (фермент тиолаза), а также реакции обратной конденсации при образовании лимонной кислоты и др. Все ферменты, участвующие в реакциях Т. к. ц., локализованы в митохондриях, причём большинство из них прочно связаны с мембранными структурами.
Образование лимонной кислоты, с превращения которой и начинается собственно Т. к. ц., является реакцией эндергонической, и её реализация возможна благодаря использованию богатой энергией связи ацетильного остагка с KoA [СН3 (О) С~SKoA]. Далее следует изомеризация лимонной кислоты в изолимонную через промежуточную стадию образования цис-аконитовой кислоты. Продуктом дальнейшего превращения изолимонной кислоты под влиянием соответствующей дегидрогеназы является, по-видимому, щавелевоянтарная кислота, декарбоксилирование которой (вторая молекула CO2 ) приводит к a-кетоглутаровой кислоте. Кетоглутаратдегидрогеназа по ряду характеристик (высокая молекулярная масса, сложная многокомпонентная структура, ступенчатые реакции, частично те же коферменты и т.д.) напоминает действие пируватдегидрогеназы. Продуктами реакции являются CO2 (третье декарбоксилирование), НАДН×Н+ и сукцинил-КоА. На этой стадии включается сукцинил-КоА-синтетаза, катализирующая обратимую реакцию образования свободного сукцината: Сукцинил-КоА + Рнеорг. + ГДФ Û Сукцинат + KoA + ГТФ. При этой реакции осуществляется так называемое субстратное фосфорилирование, то есть образование богатого энергией гуанозинтрифосфата (ГТФ) или аденозинтрифосфата (АТФ) за счёт гуанозиндифосфата (ГДФ) и минерального фосфата (Р) с использованием энергии сукцинил-КоА. После образования сукцината вступает в действие сукцинатдегидрогеназа — флавопротеид, приводящий к фумаровой кислоте. Фумараза обеспечивает равновесие между фумаровой кислотой и яблочной, а дегидрогеназа яблочной кислоты (кофермент — НАД+) приводит к завершению Т. к. ц., то есть к образованию щавелевоуксусной кислоты. На этой стадии повторяется реакция конденсации (конденсирующий фермент) между щавелевоуксусной кислотой и ацетил-КоА, приводящая к образованию лимонной кислоты.
Энергетическая эффективность рассмотренных процессов невелика. Однако образующиеся при окислении пирувата и последующих реакциях Т. к. ц. 4 моля НАДН, 1 моль ФАДН2 и 3 моля CO2 являются важными продуктами окислительных превращений. Особенно это касается восстановленных форм НАД и ФАД. Дальнейшее их окисление осуществляется ферментами дыхательной цепи и сопряжено с фосфорилированием, то есть образованием АТФ за счёт этерификации минерального фосфата (см. Окислительное фосфорилирование ). На каждую полностью окисленную до CO2 и H2 O молекулу пирувата приходится образование не менее 15 богатых энергией фосфатных связей. Процесс окисления НАДН и ФАДН2 ферментами дыхательной цепи энергетически весьма эффективен, происходит с использованием кислорода воздуха, приводит к образованию воды и служит основным источником энергетических ресурсов клетки. Однако в его непосредственной реализации ферменты Т. к. ц. не участвуют. См. также Окисление биологическое , Тканевое дыхание .
Лит.: Кребс Г., Корнберг Г., Превращения энергии в живых системах, пер. с англ., М., 1959; Филиппович Ю. Б., Основы биохимии, М., 1969; Ленинджер А., Биохимия, пер. с англ., М., 1974, гл. 16.
С. Е. Северин.
Схема цикла трикарбоновых кислот. В рамках — ферменты и окислённые формы коферментов, в двойных рамках — восстановленные коферменты и CO2 .
Трикветрум
Трикве'трум (от лат. Triquetrus — треугольный), трикветр, линейка параллактическая, древний астрономический угломерный инструмент, применявшийся для измерения зенитных расстояний небесных светил. Состоял из трёх шарнирно-соединённых стержней, образующих равнобедренный треугольник, у которого угол при вершине мог изменяться в соответствии с измеряемым зенитным расстоянием. Мерой угла служила длина стержня, находившегося в основании треугольника. Т. использовался при астрономических наблюдениях вплоть до 16 в. (Н. Коперник ).