Достаточно наладить нормальное легочное дыхание (путем изменения объема грудной полости), как приведенные выше цепочки начинают «раскручиваться» в обратную сторону. Убрав «подчелюстной насос», мы можем сделать череп высоким и узким, подвести жевательную мускулатуру к челюсти не сзади, а сверху (как у нас с вами), уменьшить – за счет улучшения рычага – ее объем и отдать освободившееся место под «мозги»; кроме того, такое расположение жевательных мышц позволит в дальнейшем не только удерживать схваченную добычу, но и пережевывать пищу (рисунок 37). Освободив кожу от дыхательных функций, мы получаем возможность разделить круги кровообращения и резко интенсифицировать обмен веществ. Эти инженерные решения вполне однозначны, равно как и необходимость одеть икринку особой оболочкой – амнионом, как бы создающей для зародыша маленький искусственный водоем и делающим его развитие независимым от водной среды; отсюда фундаментальное разделение позвоночных на «прикованных к воде» анамний (рыб и амфибий), и «истинно сухопутных» амниот – рептилий, птиц и млекопитающих). А вот дальше начинается интереснейший эволюционный выбор, когда вариантов – два, в обоих есть свои плюсы и свои минусы.
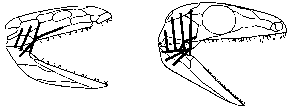
РИСУНОК 37. Расположение жевательной мускулатуры лабиринтодонта Ichthyostega (а) и примитивной капториноморфной рептилии Paleothyrus (б).
Главная проблема, которую нам предстоит решать теперь, когда осуществлен настоящий выход на сушу – это экономия воды. Возникает вопрос: как нам быть с кожей, доставшейся в наследство от амфибий – мягкой и влажной, богатой железами? Можно ее полностью заизолировать, создав на поверхности водонепроницаемый роговой слой – а можно лишь модифицировать, сохранив ее основные характеристики; оба решения вполне реализуемы, и каждое из них влечет за собою целую цепь физиологических следствий.
Создав сухую кожу с роговым покрытием, мы сводим к минимуму потери влаги: организм становится практически независимым от внешних источников воды (как одетые в «пустынные скафандры» герои фантастического романа «Дюна»). Однако за удобство надо платить.
Во-первых, необходимо перестроить выделительную систему. Почки амфибий функционально не отличаются от рыбьих и предназначены для выведения из организма избытка воды (которую тот постоянно «насасывает» за счет осмоса из гипотонической внешней среды). Проблема удаления конечного продукта белкового обмена – весьма токсичной мочевины CO(NH2)2 – решается в этом случае элементарно: ее просто растворяют в водяном потоке, который так и так постоянно «течет сквозь организм». Иное дело – когда мы начинаем экономить воду, потребляя ее по минимуму; легко создать канализацию в населенном пункте, стоящем на реке, а что делать, если в твоем распоряжении лишь артезианский колодец? Тогда «почки выведения» необходимо заменить на «почки сбережения», призванные выводить во внешнюю среду как можно меньше воды. При этом приходится менять конечный продукт белкового обмена с мочевины на менее токсичную мочевую кислоту – а этот дополнительный «технологический цикл» весьма энергоемок[52] .
Во-вторых (и это даже более важно), сухая, лишенная желез кожа создает большие трудности с терморегуляцией – а при жизни на суше, где обычны резкие температурные перепады, эта проблема одна из важнейших. При голых, ороговевших покровах равно затруднены и теплоизоляция (в холод), и теплосброс (в жару) – потому для этой эволюционной линии путь к достижению гомойотермии (поддержанию постоянной, независимой от внешней среды температуры тела) если и не закрыт полностью, то очень сильно затруднен.
Если же мы решим не гнаться за экономией воды и оставим кожу железистой, не ороговевшей, то перед нами откроется множество новых возможностей. Эти кожные железы можно преобразовать в самые разнообразные структуры. Можно превратить их в волоски, выполняющие осязательные функции, а сделав эти волоски достаточно густыми, создать теплоизолирующий покров – шерсть. Можно превратить их дополнительный орган выделения – потовые железы, которые являются еще и терморегулятором (пот, испаряясь, охлаждает поверхность тела); можно, наконец, изменить состав их выделений, превратив эти железы в млечные, и выкармливать с их помощью детенышей. Как легко видеть, в этой эволюционной линии, с ее терморегуляторными возможностями, появление гомойотермии просто-таки напрашивается – да так оно и было на самом деле. Правда, по степени зависимости от источников воды эти существа будут не слишком превосходить амфибий – но ведь за всё в этом мире приходится платить... А с другой стороны – раз уж внутренняя среда нашего организма все равно будет существовать как бы в постоянном водяном токе, нет нужды радикально перестраивать почки на «водосберегающую технологию» и возиться со сменой конечного продукта азотного обмена!
Что же это за «две эволюционные линии»? Традиционный взгляд на эволюцию рептилий – он и поныне отражен во многих учебниках – предполагает, что от каких-то антракозавров (здесь часто поминают сеймурию, имевшую уже рептилийное строение челюсти и грудной клетки, пригодной для реберного дыхания, но еще не расставшуюся с водой – личинки ее так и оставались «головастиками» с наружными жабрами) происходит некий гипотетический «общий предок пресмыкающихся», который обладает уже всем набором рептилийных признаков (водонепроницаемой кожей, почками сбережения и т.д.). От него-то и берут начало основные эволюционные ветви этого класса – анапсиды, синапсиды диапсиды и эвриапсиды; этим группам зачастую придают статус подклассов. Они выделены на основе того, каким способом в каждом из них трансформировался исходный стегальный (крышевидный) череп лабиритодонтов (рисунок 38). У анапсид (сюда относятся черепахи и ряд вымерших групп, например, парейазавры) сохраняется исходный тип черепа – массивный, лишенный облегчающих конструкцию отверстий. У синапсид – «зверообразных ящеров» , чьи зубы были дифференцированы по форме и функциям (пеликозавров и более продвинутых терапсид, которых традиционно считают предками млекопитающих) возникает одно височное окно, от краев которого отходят челюстные мышцы, и возникает единственная скуловая дуга. У диапсид же височных окон возникает два, и соответственно, две скуловых дуги – верхняя и нижняя; к этой группе принадлежат большинство рептилий, как современных (ящерицы со змеями, гаттерии и крокодилы), так и вымерших (динозавры, птерозавры). К эвриапсидам относят несколько вымерших морских групп (ихтиозавров, плезиозавров и родственных им нотозавров и плакодонтов); их черепа имеют единственное височное окно, но образуется оно иным образом, чем у синапсид[53] . Необходимо заметить, что черепа млекопитающих и птиц могут быть отнесены к, соответственно, синапсидному и диапсидному типам – что отражает происхождение этих высших тетрапод.