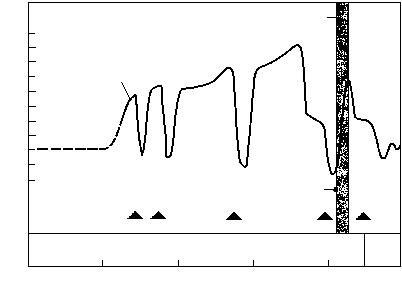
РИСУНОК 20. Ключевые характеристики истории позднего протерозоя. Изменения в ходе кривой захоронения органического углерода соответствуют ледниковым эпохам и интенсивному образованию железорудных формаций.
О том, что на Земле в это время действительно существенно возросло содержание кислорода, могут свидетельствовать и другие факты. Во-первых, в океанах вновь происходит осаждение джеспеллитов, прекратившееся около 1,8 млрд лет назад (см. рисунок 16). Это может означать, что кислород впервые насытил глубоководные части океана, располагающиеся ниже фотической зоны, и при этом произошло осаждение еще сохранявшихся в тех глубинах запасов закисного железа. Во-вторых, это было время как минимум трех покровных оледенений, последнее из которых – Лапландское (600 млн лет назад) – было, по всей видимости, самым крупным за всю историю Земли. Одной же из причин начала оледенений считают так называемый «обратный парниковый эффект»: когда в атмосфере планеты отношение О2/СО2 смещается в сторону кислорода, планета начинает гораздо хуже удерживать тепло, доставляемое центральным светилом.
Все это, вероятно, стало следствием событий, произошедших в биосфере около 1,2 млрд лет назад, когда начался постоянный рост численности и видового разнообразия эукариотного фитопланктона, продолжавщийся до 600 млн лет назад. Известно, что в планктонных экосистемах детритная пищевая цепь развита очень слабо, а оборот вещества и энергии идет почти исключительно по пастбищной цепи; практически вся мертвая органика «проваливается» из экосистемы вниз, на дно водоема, где и утилизуется в тамошних детритных цепях. Однако донные детритные цепи протерозоя существуют в условиях сильного недостатка кислорода, и потому обладают ограниченой «пропускной способностью». Начиная с некоторого момента они перестают справляться с тем нарастающим потоком органики, что поставляется сверху наращивающими свою продуктивность фитопланктонными сообществами. Другим источником органики могут быть лишенные в ту пору сколь-нибудь эффективных консументов («травоядных») водорослевые луга. В результате органический углерод начинает захораниваться в осадках на дне океана; оборотной стороной этого процесса является, как мы теперь знаем, накопление свободного кислорода, приводящее – через «обратный парниковый эффект» – к оледенению.
Сочетание этих двух факторов – заметный рост содержания кислорода и появление ледников – создает в гидросфере планеты принципиально новую ситуацию. До сих пор кислород мог распространяться из фотической зоны, где он вырабатывается, в нижележащие слои океана только за счет такого несовершенного механизма, как диффузия: ведь более богатая кислородом вода поверхностного слоя – более теплая, а потому она «не тонет». Заметим, что само «богатство» это весьма относительное, т.к. чем теплее вода, тем меньшее количество газа (при равном давлении) может быть в ней растворено. Известно, что вода имеет минимальную плотность при температуре 0ºC и ниже (лед), а максимальную – при 4ºC, поэтому с появлением на планете льда и «тяжелой» – четырехградусной – воды картина меняется. Плавающий по поверхности лед охлаждает омывающую его воду, и когда ее температура достигает 4ºC, она «тонет» – вместе с растворенными в ней газами из поверхностного слоя; в океане образуется холодный придонный слой – психросфера. Таким образом, в гидросфере возникает «ленточный транспортер», доставляющий кислород в ее глубокие слои (одним из следствий этого процесса стало осаждение в океанах джеспеллитов). Оледенения случались на Земле и раньше (например, Гуронское оледенение 2,3-2,5 млрд лет назад), но тогда этому транспортеру еще нечего было переносить; теперь же (650-600 млн лет назад) вся гидросфера оказалась насыщенной кислородом до того уровня, который допускает существование макроскопических животных.
В 1947 г. Р. Спригг сделал в местечке Эдиакара в Южной Австралии одно из самых замечательных открытий за всю историю палеонтологии. Им была найдена и описана богатая фауна удивительных бесскелетных организмов, получившая название эдиакарской. Первоначально эту фауну сочли кембрийской, однако в 1959 г. М. Глесснер правильно датировал ее концом докембрия – вендом (620-600 млн лет назад); таким образом, период достоверного существования на Земле многоклеточных животных удлинился почти на на 100 млн лет. В дальнейшем эдиакарскую фауну нашли еще в нескольких районах мира (Намибия, Ньюфаундленд); более того, выяснилось, что этих существ находили и ранее (например, на Украине в 1916 г.), однако принимали за неорганические остатки. Самое же богатое и наиболее информативное в плане палеоэкологии местонахождение (многие тысячи особей, относящихся к нескольким десяткам видов) известно в России, на Белом море.
В то время как все многочисленные группы многоклеточных, появившиеся в начале кембрия (540 млн лет назад), были представлены мелкими организмами (миллиметры или первые сантиметры), эдиакарская фауна состояла из крупных или очень крупных беспозвоночных, размером до полутора метров. Среди них были как радиально-симметричные формы, назваемые «медузоидами», так и билатерально-симметричные; одни из них (петалонамы) внешне напоминают современные кораллы морские перья (Pennatularia), другие (дикинсония и сприггина) – кольчатых червей и членистоногих (рисунок 21). Первые исследователи эдиакарской фауны (Спригг и Глесснер) считали эти формы реальными предками современных кишечнополостных и червей и включали их в состав соответствующих типов и классов животных. Эта точка зрения имеет сторонников и поныне («австралийская школа»), однако большинство исследователей считает, что сходство тут чисто внешнее, и эдиакарские организмы представляют собой нечто совершенно особенное. А. Зейлахер (1984) ввел для этих существ специальное собирательное название – вендобионты.

РИСУНОК 21. (а) – Различные представители эдиакарской фауны – радиально– и билатеральносимметричные; (б) – строение дикинсонии.