Биохимический «маятник» человека состоит из факторов транскрипции CLOCK и BMAL1 и генов-мишеней: трех генов с говорящим названием Periods (Per 1, 2 и 3) и двух криптохромов – Cryptochromes (Cry 1 и 2). Следует отметить, что криптохромы – это очень древние и эволюционно консервативные белки. Они присутствуют не только у животных, но и у растений, где выполняют функцию сенсоров света и тьмы и таким образом регулируют также циркадные ритмы[16].
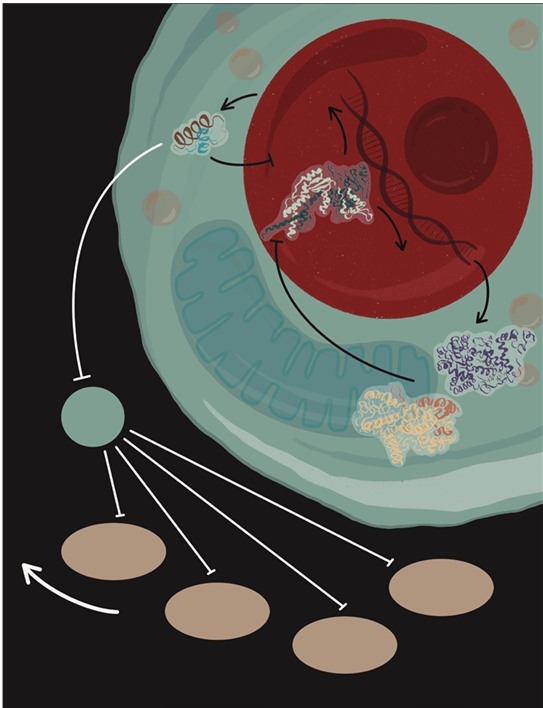
CLOCK и BMAL1 активируют PER и CRY, а те в свою очередь ингибируют транскрипционную активность BMAL1-CLOCK, формируя петлю отрицательной обратной связи. Это приводит к ритмической экспрессии комплекса BMAL1-CLOCK с периодичностью 24 часа, и этот комплекс служит своеобразным биохимическим маятником.
Рисунок 5.1. Работа комплекса CLOCK/BMAL1 лежит в основе механизма циркадных ритмов млекопитающих. Он активирует несколько генов, среди которых Per 1, 2, 3 и Cry 1, 2. Белки – продукты этих генов – перемещаются в ядро, где ингибируют транскрипционную активность комплекса CLOCK/BMAL1. REV-ERBα – еще один ген, контролируемый комплексом CLOCK/BMAL1, белковым продуктом которого является отрицательный регулятор экспрессии Bmal1. Кроме того, REV-ERBα подавляет экспрессию ингибитора клеточного цикла – p21. В отсутствие BMAL1 подавление REV-ERBα приводит к высокой экспрессии p21 и задержке зародышевых клеток волос в фазе G1, что останавливает анаген.
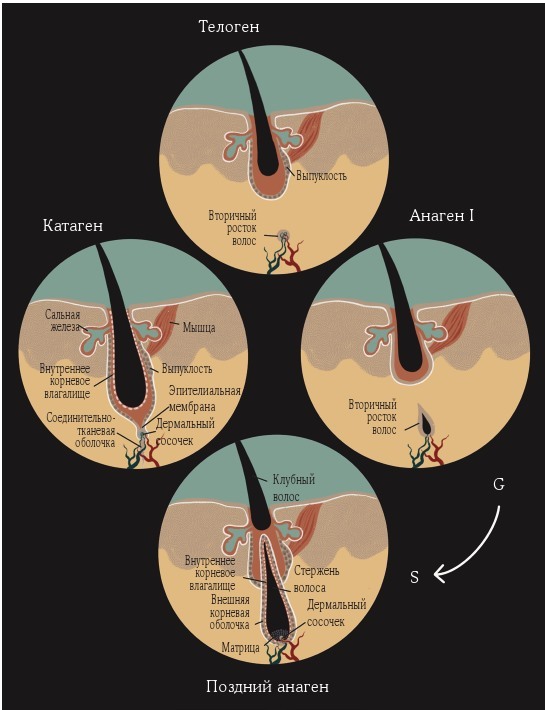
Рисунок 5.2. Обозначения: APM – мышца arrector pili, Bu – выпуклость, CH – клубный волос, CTS – соединительнотканная оболочка, DP – дермальный сосочек, EM – эпителиальная мембрана, HS – стержень волоса, IRS – внутреннее корневое влагалище, Ma – матрица, ORS – внешняя корневая оболочка, SB – сальная железа, SHG – вторичный росток волос[17].
Функции генов циркадных ритмов не исчерпываются только самими циркадными ритмами – они задействованы также и в росте волос. В телогене и раннем анагене активируются гены-мишени комплекса CLOCK/BMAL1. Из-за того что в коже они экспрессируются с четким циркадным ритмом, амплитуда уровня их экспрессии наиболее высока во время телогена и раннего анагена. Это указывает на то, что их активность зависит как от циркадного механизма, так и от цикла роста волос[18][19][20].
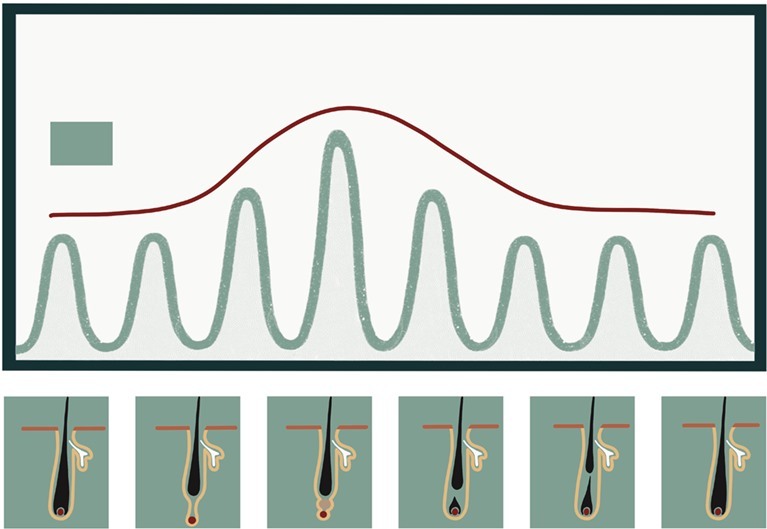
Рисунок 6. Диаграмма, отражающая ритмическую циркадную экспрессию гена Dbp (активатор транскрипции) на разных фазах цикла роста волос. Амплитуда экспрессии Dbp коррелирует с прогрессированием цикла волосяного фолликула: она становится максимальной во время телогена[21].
Гены циркадных часов экспрессируют все типы клеток кожи, но местом наиболее заметной ритмической экспрессии циркадных генов во время телогена и раннего анагена является вторичный зародыш волоса. Это отдел, расположенный между дермальным сосочком и выпуклостью. Он содержит пролиферативные стволовые клетки, которые мигрировали из выпуклости во время позднего катагена и раннего телогена[22][23][24]. Вторичные зародышевые клетки волоса во время инициации анагена активируются первичными, в результате чего формируются размножающиеся клетки волосяного матрикса, а в конечном итоге и стержень волоса[25][26][27].
У мышей, мутантных по генам Clock и Bmal1, анаген значительно задерживается, причем этот эффект наиболее выражен у мышей с дефицитом именно Bmal1. Такие мыши имеют целый ряд отличий от мышей дикого типа. Исследования показали следующее.
• В то время как в ходе анагена в большинстве волосяных фолликулов у здоровых мышей уже образовались волосяной матрикс и стержень волоса с волосяной луковицей, мутантные по Bmal1 животные все еще оставались в самой первой его фазе. В результате волосяные фолликулы экспериментальных животных возобновили нормальное развитие цикла роста волос лишь после почти недельной задержки. При этом аномалий в структуре зрелых фолликулов анагена у мутантных по Bmal1 или Clock мышей не было, что подтверждает участие генов циркадных часов, в первую очередь, в механизмах перехода телоген-анаген.
• Во вторичном зародыше волос раннего анагена в волосяных фолликулах мутантных по Bmal1 животных отсутствовали делящиеся клетки. При этом эпидермис и дерма этих мышей их содержали, значит, дефект пролиферации специфичен именно для волосяного фолликула[28][29].
• Во вторичном зародыше волос волосяных фолликулов мутантных животных отсутствовал фосфорилированный белок ретинобластомы (Rb) (маркер прохождения клеточного цикла через контрольную точку G1-S), хотя в норме его очень много. Это говорит о том, что в волосяных фолликулах экспериментальных мышей клетки-предшественники вторичного зародыша волос раннего анагена задерживаются в контрольной точке клеточного цикла G1-S.
• В коже мышей с дефицитом Bmal1 во время телогена оказалась затронута экспрессия нескольких генов-мишеней CLOCK-BMAL1, в том числе Rev-Erbα. Она снижалась в целых пятнадцать раз! Мы знаем, что REV-ERBα напрямую подавляет экспрессию гена, кодирующего ингибитор клеточного цикла G1-p21. Таким образом, в мутантной коже p21 активируется примерно в 2,5 раза сильнее.
• Активация p21 у экспериментальных животных привела к замедлению прогрессирования клеточного цикла в фазах G1-S в клетках-предшественниках вторичного зародыша волос[30].
Пигментация волос
Пигментация волос – это результат активности специальных клеток меланоцитов. Они названы так потому, что эти клетки производят пигмент меланин и откладывают его в стержень волоса по мере его формирования[31]. Меланоциты находятся в луковице волосяного фолликула.
Пигментация фолликулов чувствительна к многочисленным внутренним факторам. На нее влияют:
• фаза цикла роста волос;
• место расположения фолликула на теле;
• раса и пол;
• чувствительность к гормонам;
• генетические дефекты;
• возрастные изменения.
Пигментацию волос регулирует множество различных веществ, таких как факторы роста, цитокины, гормоны, нейропептиды и нейромедиаторы, эйкозаноиды, циклические нуклеотиды, питательные вещества, микроэлементы. Многие из этих веществ могут действовать через механизмы межклеточной сигнализации[32].
Значение пигментации
Пигментация необходима по нескольким причинам, которые являются различными для разных видов животных. Она выполняет несколько функций, основными из которых являются следующие:
• защита кожи от ультрафиолетового излучения солнца;